Download
1 / 36
370 likes | 756 Views
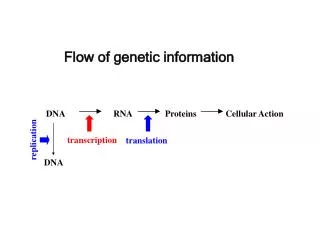
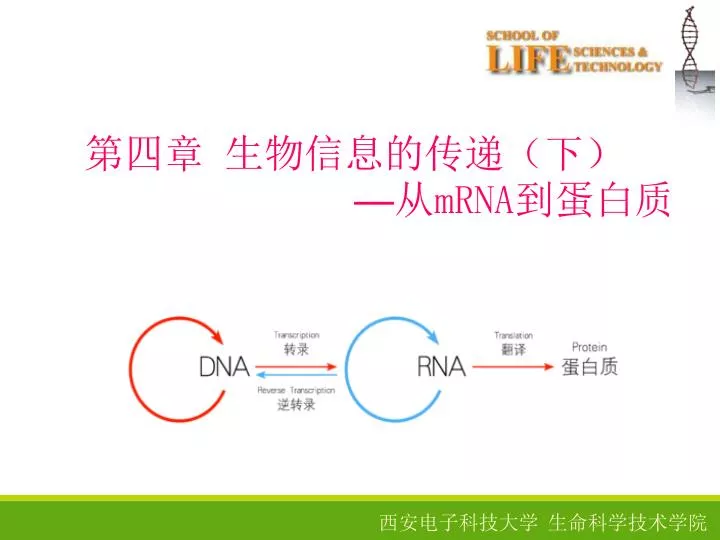
第四章 生物信息的传递(下) — 从 mRNA 到蛋白质. 3. DNA→mRNA→蛋白质. DNA的复制 解旋→引发→复制→终止 mRNA的转录 模板识别→转录起始→转录延伸→转录终止 蛋白质的合成 翻译的起始→肽链的延伸→肽链的终止及释放. 翻译: 指将 mRNA 链 上的核甘酸从一个特定的起始位点开始,按每 三个核甘酸 代表一个氨基酸的原则,依次合成一条多肽链的过程。. 蛋白质合成的场所是 蛋白质合成的模板是 模板与氨基酸之间的接合体是 蛋白质合成的原料是. 核糖体. mRNA. tRNA. 20 种氨基酸.

E N D
第四章 生物信息的传递(下)—从mRNA到蛋白质
3 DNA→mRNA→蛋白质 DNA的复制 解旋→引发→复制→终止 mRNA的转录 模板识别→转录起始→转录延伸→转录终止 蛋白质的合成 翻译的起始→肽链的延伸→肽链的终止及释放
翻译:指将mRNA链上的核甘酸从一个特定的起始位点开始,按每三个核甘酸代表一个氨基酸的原则,依次合成一条多肽链的过程。 蛋白质合成的场所是 蛋白质合成的模板是 模板与氨基酸之间的接合体是 蛋白质合成的原料是 核糖体 mRNA tRNA 20种氨基酸
● 遗传密码——三联子 ● tRNA的结构、功能与种类 ● 核糖体的结构与功能 ● 蛋白质合成的过程 ● 蛋白质的运转机制 Contents
7 三联子密码的来源 理论预测 4种核苷酸20种天然氨基酸 实验证明 插入或删除一个碱基,会改变最终的全部氨基酸序 列 同时插入和删除实验,除突变密码子所对应的氨基 酸,其它氨基酸序列都不变 同时删除三个碱基,翻译产生少了一个氨基酸的蛋 白质
(一)三联子密码定义 mRNA链上每三个核甘酸翻译成蛋白质多肽链上的一个氨基酸,这三个核甘酸就称为密码子或三联子密码(triplet codon)。 一、遗传密码——三联子 mRNA 5’ GCU AGU ACA AAA CCU 3’
(二)三联子密码破译 AUCGACCUGAGC mRNA 3′ 5′ 4 (×) 20 mRNA AUCGACCUGAGC 3′ 5′ 42=16 20 (×) mRNA AUCGACCUGAGC 3′ 5′ 43=64 20 (√) 核甘酸序列 氨基酸序列
遗传密码的破译,即确定代表每种 氨基酸的具体密码。 ●至1966年,20种氨基酸对应的61个密码子和三个终止密码子全部被查清。
13 (三)遗传密码的性质 连续性:连续地读码 简并性:几个密码子编码一种氨基酸 通用性和特殊性:对不同物种通用 摆动性:密码子与反密码子: mRNA与tRNA的结合
(三)遗传密码的性质 1、 连续性 编码蛋白质氨基酸序列的各个三联体密码连续阅读,密码间既无间断也无交叉。
从mRNA 5端起始密码子AUG到3端终止密码子之间的核苷酸序列,各个三联体密码子连续排列编码一个蛋白质多肽链,称为开放阅读框架(open reading frame,ORF)。
(三)遗传密码的性质 2、简并性 由一种以上密码子编码同一个氨基酸的现象称为简并(degeneracy),对应于同一氨基酸的密码子称为同义密码子(synonymous codon)。
许多氨基酸都有多个密码子 甲硫氨酸 色氨酸
编码某一氨基酸的密码子越多,该氨基酸在蛋白质中出现的频率就越高。Arg例外编码某一氨基酸的密码子越多,该氨基酸在蛋白质中出现的频率就越高。Arg例外
(三)遗传密码的性质 3、通用性与特殊性 • 蛋白质生物合成的整套密码,从原核生物到人类都通用。 • 已发现少数例外,如动物细胞的线粒体、植物细胞的叶绿体。
(三)遗传密码的性质 4、摆动性 转运氨基酸的tRNA上的反密码子需要通过碱基互补与mRNA上的遗传密码子反向配对结合,在密码子与反密码子的配对中,前两对严格遵守碱基配对原则,第三对碱基有一定的自由度,可以“摆动”,这种现象称为密码子的摆动性。
17 密码子与反密码子摆动配对 tRNA 321 123 密码子 321 123 密码子 反密码子 反密码子 mRNA 反密码子第一位的U可与A或G匹配 摆动假说:密码子和反密码子的前两对严格遵守碱 基配对原则,第三对碱基有一定自由度
摆动假说分析tRNA能识别几个密码子由反密码子的第一个碱基的性质决定摆动假说分析tRNA能识别几个密码子由反密码子的第一个碱基的性质决定
tRNA 反密码子第一位是I(次黄嘌呤)时,密码子第三位可以是U、C、A
● 遗传密码——三联子 ● tRNA的结构、功能与种类 ● 核糖体的结构与功能 ● 蛋白质合成的过程 ● 蛋白质的运转机制 Contents
二、tRNA的结构、功能与种类 tRNA在蛋白质合成中的地位 第二遗传密码 共同特征 存在经过特殊修饰的碱基 3’端都以CCA-OH结束,是tRNA与相应氨基 酸结合的位点
21 (1)三叶草型二级结构 受体臂 TΨC臂 多余臂 D臂 反密码子臂
三级结构:“L”形 螺旋1 螺旋2 氢键维持
连接氨基酸 L型结构将分子中 两个不同功能基团 最大限度分离 连接mRNA
1、解读mRNA的遗传信息 2、运输的工具,运载氨基酸 (二) tRNA的功能 tRNA有两个关键部位: ● 3’端CCA:接受氨基酸,形成氨酰-tRNA。 ● 与mRNA结合部位—反密码子部位
tRNA凭借自身的反密码子与mRNA链上的密码子相识别,把所带氨基酸放到肽链的一定位置。tRNA凭借自身的反密码子与mRNA链上的密码子相识别,把所带氨基酸放到肽链的一定位置。 CCA-OH CCA-OH 3’ 5’ C C G I 5’ 3’ G G C
1、起始tRNA和延伸tRNA (三) tRNA的种类 能特异地识别mRNA模板上起始密码子的tRNA称 起始tRNA,其他tRNA统称为延伸tRNA。
真核生物:起始密码子AUG 所编码的氨基酸是甲硫氨酸Met,起始AA-tRNA为Met-tRNAMet。 原核生物:起始密码子AUG 所编码的氨基酸并不是 甲硫氨酸本身, 而是甲酰甲硫氨酸,起始AA-tRNA为fMet-tRNAfMet
2、同工tRNA (三) tRNA的种类 • 代表同一种氨基酸的tRNA称为同工tRNA。 • 同工tRNA要有不同的反密码子以识别该氨基酸的各种同义密码子 • 某种结构上的共同性,能被相同的氨基酰-tRNA合成酶识别。
无义突变:在蛋白质的结构基因中,一个核苷酸的改变可能使代表某个氨基酸的密码子变成终止密码子(UAG、UGA、UAA),使蛋白质合成提前终止,合成无功能的或无意义的多肽,这种突变就称为无义突变。 错义突变:由于结构基因中某个核甘酸的变化使一种氨基酸的密码子变为另一种氨基酸的密码子,这种基因突变叫错义突变。 GGA(甘氨酸) AGA(精氨酸) 3、校正tRNA (三) tRNA的种类
无义突变抑制 酪氨酸 校正tRNA与释放因子竞争识别密码子,抑制终止密码子,造成翻译过程的通读。
错义突变抑制 甘氨酸 校正tRNA与正常tRNA竞争,校正错义突变
33 氨酰-tRNA合成酶 AA-tRNA合成酶催化氨基酸与tRNA特异 性结合 氨酰-tRNA合成酶 AA+tRNA+ATP AA-tRNA+AMP+PPi 氨基酸活化 AA+ATP+酶(E)→E-AA-AMP+PPi E-AA-AMP+tRNA→AA-tRNA+E+AMP氨酰基转移