Download

1 / 79
830 likes | 1.05k Views
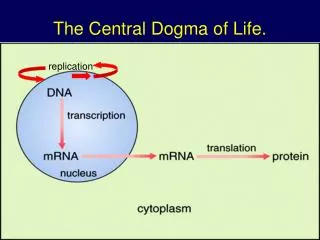
第十七章 蛋白质的生物合成 与修饰 Protein Synthesis and modification. 第一节 概述. 主讲老师:华南师范大学生命科学学院 陈文利. 翻 译. 生物体合成 mRNA 后, mRNA 中的遗传信息转变成为蛋白质中氨基酸排序的过程称为 翻译 。. 条件:. 1、 原料 20种氨基酸. 2、 能量 ATP 和 GTP. 3、 催化剂: 酶、蛋白质因子、无机离子( Mg + ,K + ). 4、 运载工具: tRNA. 5、 模板: mRNA( 每个 A.A 由三个碱基确定).

E N D
第十七章 蛋白质的生物合成与修饰 Protein Synthesis and modification 第一节 概述 主讲老师:华南师范大学生命科学学院 陈文利
翻 译 生物体合成mRNA后,mRNA中的遗传信息转变成为蛋白质中氨基酸排序的过程称为翻译。 条件: 1、原料 20种氨基酸 2、能量 ATP和GTP 3、催化剂: 酶、蛋白质因子、无机离子(Mg+,K + ) 4、运载工具:tRNA 5、模板:mRNA(每个A.A由三个碱基确定) 6、装配机:核糖体(rRNA、蛋白质)
第三节 遗传密码的几个重要特性 遗传密码: mRNA分子上从5’3’的方向,每三个碱基形成的三联体,组成一个遗传密码子(codon)。
遗传密码表:共有64个密码子,其中61 个是编码氨基酸。其中AUG ↗起始密码子 ↘Met的密码子。 有三个密码子是终止密码子:UAA、UAG、UGA, 这三组密码子不能被tRNA阅读,只能被 肽链释放因子识别。
遗传密码的基本特点(5个性): 1、密码子的简并性 2、密码子的连续性 3、密码子的不重叠性 4、密码子的摆动性 5、密码子的通用性 6、特殊密码子 返回
1、密码子的简并性 一个氨基酸具有多个密码子的现象称为密码子的简并性(degeneracy)。 这些编码同一种氨基酸的多个密码子称为同义密码子(synonymous codon) 遗传密码的基本特点
2、密码子的连续性 要正确阅读密 码必须按一定的密码框架(reading frame) 从一个正确的起点开始,一个不漏地挨 着读下去,直到碰到终止信号为止。 遗传密码的基本特点
3、密码子的不重叠性(non-overlapping) 绝大多数生物中读码规则是不重叠的。少数大肠杆菌噬菌体的RNA基因组中,部分基因的遗传密码却是重叠的。 遗传密码的基本特点
4、密码子的摆动性 P423 密码子的第一、第二位专一性很强,第三位专一性就弱。 已证明,密码子的专一性主要由头两位碱基决定,Crick对第三位碱基的这一特性给于一个专门的术语,称“摆动性” wobble. 遗传密码的基本特点
tRNAs • Secondary & Tertiary structure • Isoacceptor tRNAs (同工受体tRNA) Different tRNAs carrying the same aa • tRNA identity- “2nd genetic code” P431 Special sequence elements on tRNAs that can be recognized by aaRS and then determine which aa is charged Positive elements & Negative elements • Some special tRNAs 1) Initiator tRNA: tRNAfMet & tRNAiMet 2) Suppressor tRNA(校正tRNA) 3) tmRNA
5、密码子的通用性 遗传密码表属于完全通用 。 返回 遗传密码的基本特点
General features • With mRNA as template, tRNA as carrier for aa, ribosome as assembly site • Translational polarity 1) Extends in N-end → C-end how to prove? 2) Reads mRNAs in 5’ → 3’ • Triplet codon 1) How many bases determine one aa? “three determine one” 2) Cracking of the genetic code 3) Features of the genetic code • Ribosomes recognize aa-tRNA just by virtue of the base-pairing interaction between codons and anticodons • Wobble hypothesis
The Nature of the Genetic Code • All the codons have meaning: 61 specify amino acids, and the other 3 are "nonsense" or "stop" codons • The code is unambiguous - only one amino acid is indicated by each of the 61 codons (wobble密码子的摆动性) • The code is degenerate - except for Trp and Met, each amino acid is coded by two or more codons (密码子的简并性) • Codons representing the same or similar amino acids are similar in sequence
2nd base pyrimidine: usually nonpolar amino acid 2nd base purine: usually polar or charged aa The code is not overlapping(密码子的不重叠性) The base sequence is read from a fixed starting point, with no punctuation(密码子的连续性) Universal & Unusual
第四节 蛋白质的生物合成 一、核糖体(ribosome)是一个复杂的超分子结构 核糖体可以看作是一个大分子的机构,它具有许多精密的配合部分,来挑选并管理参与蛋白质合成的各个组分。它参与多肽链的启动,延长和终止的各种因子的识别。
二、核糖体(ribosome) 原核生物核糖体 5S rRNA, 23S rRNA 50S 34种蛋白质 70S 16S rRNA 30S 21种蛋白质
真核生物核糖体 5SrRNA,5.8SrRNA,28SrRNA 60S 49种蛋白质 80S 18SrRNA 40S 33种蛋白质
二、转移RNA具有特征性结构 tRNA是氨基酸的搬运工具。 氨基酸的同工受体:能够运输同一种氨基酸的多种tRNA分子。 tRNA分子上与蛋白质生物合成有关的位 点至少有四个: 1)3端—CCA上的氨基酸接受位点 2)识别氨酰—tRNA合成酶的位点 3)核糖体识别位点,使延长中的肽链 附着于核糖体上 4)反密码子位点
Functional sites on Ribosome • Three sites for tRNA 1) The A (acceptor) site - where aa-tRNAs come in (except the first one) and where peptidyl-tRNA is after peptide bond formation and before translocation. 2) The P (peptidyl-tRNA) site - where peptidyl-tRNA is before peptide bond formation. 3) The E (exit) site - where the uncharged tRNA from the P site goes after translocation. • Peptidyl transferase - the active site that catalyzes formation of the peptide bond (23S rRNA in Prokaryotes) • Polypeptide exit channel • mRNA binding site
三、氨酰-tRNA合成酶和它们催化的反应 P429 四、一些氨酰-tRNA合成酶具有校对功能 氨酰—tRNA合成酶 , 酶的专一性表现在: a)识别氨基酸 b)识别tRNA(倒L型的三级结构) c)进行二次核对作用
E.Coli蛋白质的生物合成: 1.氨基酸的活化: 由高度特异的氨酰-tRNA合成酶 (aminoacyl-tRNA synthetase)催化,反应分两步 总反应式
五、氨酰-tRNA合成酶对tRNA的识别 六、多肽链的合成从氨基末端开始
七、一个特定氨基酸起始蛋白质的合成 起始氨酰-tRNA的形成(P432) 现已清楚,原核细胞中多肽的合成都有 自甲硫氨酸开始,但并不是以甲硫氨酰-tRNA作起始物,而是以N-甲酰甲硫氨酰-tRNA的形式起始。
在E.Coli和其它原核生物中与这起始密码(AUG)相对应的tRNA是甲酰甲硫氨酰—tRNA(fMet-tRNAfMet),这是起始tRNA。在E.Coli和其它原核生物中与这起始密码(AUG)相对应的tRNA是甲酰甲硫氨酰—tRNA(fMet-tRNAfMet),这是起始tRNA。
起始tRNA怎样形成?由甲硫氨酰-tRNA甲酰化 甲酰基 甲酰基转移酶 tRNA Met-tRNA Met N10-甲酰FH4 FH4 (fMet-tRNAf)
Structure of N-formyl-methionyl-tRNA[Met] Differences with other tRNAs
细胞内有2种可携带Met的tRNA,它们都识别同样的AUG密码子,但它们的一级结构和功能不同。细胞内有2种可携带Met的tRNA,它们都识别同样的AUG密码子,但它们的一级结构和功能不同。 (1)tRNAfMet带上Met后能甲酰化,是起始tRNA,用于肽链合成的起始。 (2)tRNAmMet带上Met后不能甲酰化,用于肽链的内部,在肽链延伸中起作用。 所以AUG和GUG是兼职密码子,它们既可以作为起始密码子,作为肽链合成的起始信号,这时与之对应的氨基酸是甲酰Met。另外也可作肽链内部相应aa的密码,这时AUG编码Met,GUG编码val。
八、多肽合成起始的三个步骤 1、30S-mRNA复合物的形成 此反应须起始因子3(IF3) P位:肽基的结合部位 A位:氨酰基的结合部位 起始密码定位在P位上 SD序列 Shine-Dalgarno发现在AUG的前 方有一段富嘌呤AG区,与30S中的16SrRNA富含嘧啶的区结合
mRNAs Prokaryotic mRNAs Usually polycistronic Eukaryotic mRNAs Usually monocistronic
2.肽链的起始: • mRNA中的起始密码是AUG,少数是GUG。 • 起始密码子的上游约10个核苷酸的地方往往有一段富含嘌呤的序列称SD序列(Shine-Dalgarno序列),一般为3~10个核苷酸,它与核糖体16srRNA 3ˊ端的核苷酸序列互补,可促使核糖体与mRNA的结合。
2、30S预起始复合物(IF1,IF2,GTP) 消耗一个高能键 IF1起协助作用,促进IF3,IF2的作用,然后在IF1参与下,mRNA-30S-IF3进一步与fMet-tRNAf,GTP相结合,并释 放出IF3形成 一个30S起始复合物, 30S核糖体—mRNA-fMet-tRNAf。 肽键的合成
SD sequence inactive 70S ribosome 30S initiation complex
GDP + Pi 70S initiation complex
3. 肽链的延长: 肽链的延长需要一些延长因子或称延伸因子(elongation factor,EF) 原核生物的EF有EF-T和EF-G两类: • EF-T:延伸因子“T”,开始误认为这类延伸因子有肽基转移酶的活性peptidyl transferase,所以取转移酶的第一个字母“T” 。 • EF-G:这类延伸因子与核糖体结合时需要GTP,当GTP水解时,这类延伸因子就从核糖体上解离下来,总之涉及GTP,所以用GTP的第一个字母“G”,EF-G。
EF-T是由Tu和Ts两个亚基组成的二聚体, • EF-Ts:s是stable的意思,是稳定蛋白质, • EF-Tu: u是unstable,不稳定蛋白质。 • EF-Tu直接参加了氨酰-tRNA与核糖体的结合。
肽链的延长包括3步: • 氨酰-tRNA进入核糖体的A位 • 肽键的形成 • 移位
(1)氨酰-tRNA进入核糖体的A位 这是个比较复杂的过程,需Ts、Tu,还要 消 耗GTP。EF-Tu与GTP和氨酰-tNRA首先形成三元 复合物,才能进入A位。
(2)肽键的形成 通过一个转肽作用(transpeptidation),由肽酰转移酶(peptidyl transferase)催化,使一个酯键变成了肽键,肽基转移酶的活性由核糖体大亚基的23s rRNA承担(肽基转移酶是一种ribozyme)。嘌呤霉素对蛋白质的抑制作用就发生在肽键形成这一步。