Download

1 / 39
400 likes | 578 Views
Understanding the function of conserved non-coding regions in the human genome. Sofie Salama – Haussler lab CS273A, November 17, 2008. Haussler Lab. Dry lab – comparative genomics research Browser staff – UCSC genome browser, ENCODE data coordination center, 1000 genomes

E N D
Understanding the function of conserved non-coding regions in the human genome Sofie Salama – Haussler lab CS273A, November 17, 2008
Haussler Lab • Dry lab – comparative genomics research • Browser staff – UCSC genome browser, ENCODE data coordination center, 1000 genomes • Wet lab - Experimental analysis of interesting human genomic regions
Understanding the function of conserved non-coding regions in the human genome • Origin of conserved non-coding regions and co-regulated gene networks • Function of ultraconserved elements • Discovery of novel non-coding RNA genes • Detailed analysis of Human Accelerated Regions (HAR’s)
How are we different from chimps? • Brain anatomy • 3X larger, especially cortex • More later developing neurons of the upper cortical layers projecting within the cortex • functional asymmetries • What are the genotypic differences responsible for these phenotypic differences? Hill, R. S. & Walsh, C. A. Nature 437, 64–67 (2005)
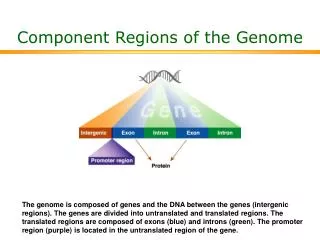
Clues from comparative genomics • Human vs. chimpanzee genome • Genomes are almost identical • BUT, almost 29 million differences • What are the important differences??? • Multiple mammalian genomes sequenced • Conservation used to identify functional elements • only 1/3 of conserved regions are protein coding
The HAR screen • Identify previously conserved regions • ≥100 bp 96% identical between the chimpananzee, mouse and rat genomes • ~35,000 mammalian conserved regions • Compare to human sequence to identify Human Accelerated Regions • Look for orthologous segments with a large number of changes • Develop statistical methods to rank and evaluate each HAR • Identified 49 regions with a significant increased substitution rate in humans (genome wide FDR<5%) Katie Pollard
HAR population resequencing Analysis of HAR1 Characterization of HAR2 knockout and knockin mice Wet lab HAR projects
Why resequence the HARs? • Positive selection • Beneficial mutation enters population • Spreads. Nearby (neutral) alleles from mutated chromosome hitchhike towards fixation – a selective sweep • Skew DAF spectrum towards both ends • Confounding factor: time • Neutral drift removes variation in 4Neff generations (~1 MYr in human) • Human/chimp ancestor 5-7 MYA Stringer Nature 2003 Noonan et al. Science 2006
Resequence HARs 1 to 49 40kb around each HAR (~2.5Mb total with 13 control regions) 24 samples (48 chromosomes) YRI hapmap samples (panel P2 Seattle SNPs) Enough to do population genetic analysis on a HAR-by-HAR basis (not like our paper on ultras in the average) High throughput sequencing technology enables cost effective investigation. Sol Katzman
“Next-Gen” Sequencing ABI SOLiD (fluoro seq by repeated ligation) 35bp reads (fragment, not mate-pair) $3-4K per run 2 slides per run multiple samples per slide barcoded samples Isolated drops on a slide 50 to 100 Million reads per slide Total 2.5Gb of reads 50% mapped? 50% enriched? 250X coverage of 2.5Mb target regions? Divide by number of samples in run for sample coverage From 1000 Genomes project: Need 11X to get both alleles @ 99% prob Need 27X average to get 11X @ 99% prob
Project Overview (part 1 of 2) Sol Katzman to Part 2
Project Overview (part 2 of 2) from Part 1 Sol Katzman
HAR population resequencing Analysis of HAR1 Characterization of HAR2 knockout and knockin mice Wet lab HAR projects
and the winner is….HAR1! • 118 bp segment with 18 changes between the human and chimp sequences
HAR1 genomic landscape • Browser gazing suggested the HAR1 element may be expressed in both orientations • rt-PCR on human tissue RNA preps suggested brain specific expression of the HAR1 element • Used RACE to clone both forward and reverse transcripts from cortical and cerebellar RNA
HAR1 is transcribed HAR1 • HAR1F expressed in brain (cerebellum, forebrain structures), ovary and testes (~1/10 of brain expression) • HAR1R expressed in brain (1/10 of HAR1F) and testes • Outside HAR1 element, little conservation beyond primates
RNA in situ hybridization superfly.ucsd.edu • Fix tissue (whole embryo or sections) • Synthesize digoxygenin labelled probe anti-sense to desired target • Hybridize, wash, visualize using enzyme linked anti-DIG anitbody
HAR1F is expressed in the in the neocortex 500µm 250µm Nelle Lambert, Marie-Alexandra Lambot, Sandra Coppens, Pierre Vanderhaeghen
Reelin and cortical development Amadio, JP & Walsh, CA, Cell 126:1033-1035 (2006)
HAR1F is expressed in the marginal zone and the cortical plate 125 µm Nelle Lambert, Marie-Alexandra Lambot, Sandra Coppens, Pierre Vanderhaeghen
Expression of HAR1F in the neocortex continues though 19 GW 1000 µm 250 µm Nelle Lambert, Marie-Alexandra Lambot, Sandra Coppens, Pierre Vanderhaeghen
Co-expression of Reelin and HAR1F in Cajal-Retzius neurons 250 µm 250 µm Nelle Lambert, Marie-Alexandra Lambot, Sandra Coppens, Pierre Vanderhaeghen
Expression of HAR1F elsewhere in the brain at later embryonic stages Nelle Lambert, Marie-Alexandra Lambot, Sandra Coppens, Pierre Vanderhaeghen
The HAR1F neocortical expression pattern is found in macaque • Expression pattern conserved since the divergence of hominoids and old world monkeys 25 MYA Colette Dehay, Pierre Vanderhaeghen
HAR1F is predicted to form a stable RNA structure Jakob Pederson
U U G G C C A A - - 0 0 10 10 30 30 Human Chimp Human - 40 - 50 - 60 Chimp - 70 DMS DMS Structure probing reveals differences in the human and chimp structures Haller Igel, Manny Ares
Resequencing/population genetics • Samples • 24 member human diversity panel (HAR1 element) • 70 Caucasian and African American (6.5 kb region) • Other primates (gorilla, orangutan, macaque) • Findings • human-specific changes fixed in the populations (NO SNPs!) • Changes happened at least 1 MYA, no evidence of a recent selective sweep • Large number of human changes extends throughout HAR1F 1st exon Sol Katzman, Bryan King, Andy Kern
Summary • HAR1 is the most extreme of a set of genomic regions showing increased substitutions specifically in the human lineage • HAR1 overlaps 2 divergent ncRNA genes, HAR1F and HAR1R • HAR1F is expressed in the neocortex in reelin producing Cajal-Retzius neurons which are critical for creating the architecture of the human cortex and also in other structures patterned by the reelin pathway • HAR1F forms a stable RNA structure and the human substitutions appear to alter this structure
What does HAR1 do??? • What is the cellular role of HAR1 ncRNAs? • Where are they localize? • Who do they interact with? • What is their role in neural development? • How do human HAR1 ncRNAs differ from other mammalian HAR1 ncRNAs?
HAR population resequencing Analysis of HAR1 Characterization of HAR2 knockout and knockin mice Wet lab HAR projects
HAR2 • 12 human substitutions in a 119 bp segment • highly conserved in amiotes, present in frog • Not in a mature transcript, no RNA secondary structure
HAR2 Genomic Neighborhood • HAR2 located in an intron of Centaurin-gamma 2 • Closest neighbor is Gastrulation and brain-specific homeobox protein 2 • CENTG2-HAR2-GBX2 relationship conserved back to frog-human ancestor
Transgenic assay for enhancer activity HAR2 Minimal Promoter LacZ Harvest at embryonic timepoints. Stain to visualize lacZ activity. How does LacZ expression compare with that of nearby genes (centg2 and gbx2)?
HAR2 is a neural-specific enhancer Bryan King and Armen Shamamian
HAR2 is a limb specific enhancer • Human HAR2 shows significant activity in the limb buds • Human HAR2 is stronger and shows a broader pattern of expression • Making the human substitutions in the chimp construct is sufficient for increased limb bud staining Prabhakar et al. (2008) Science
HAR2 targeted mutants • HAR2 knockout – marked allele is made, breeding with constitutive cre mouse to remove vector/marker sequences • HAR2 knockin human HAR2 – Have ES cell line, no chimeras yet • HAR2 knockin mouse HAR2 – Have construct Robert Sellers, Armen Shamamian
Acknowledgements Haussler Lab UCSC Collaborators Manny Ares Haller Igel Harry Noller David Feldheim Jena Yamada Nader Pourmand Pierre Vanderhaegen – Univ. of Brussels Katie Pollard – UCDUCSF/Gladstone Andy Kern - Dartmouth Funding Jeff Long, Ting Wang, Danielle Gomez HHMI, NIDA