Download

1 / 45
450 likes | 610 Views
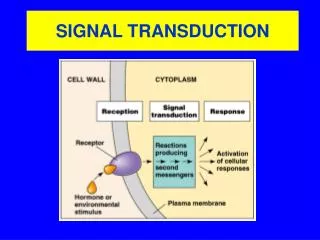
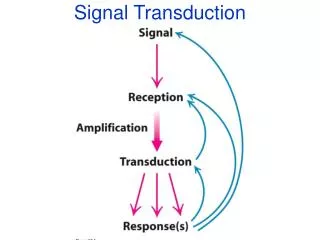
Calcium Signal Transduction. By what mechanisms are calcium signals read and translated into biochemical response? What is the structural basis for the function of the proteins involved? How is the specificity of different signalling pathways generated?.

E N D
Calcium Signal Transduction By what mechanisms are calcium signals read and translated into biochemical response?What is the structural basis for the function of the proteins involved?How is the specificity of different signalling pathways generated?
Now we understand how the calcium signal is read and transduced into biochemical responseThe next step is to examine how the diversity in the functions of EF-hand proteins is achieved.
EF-hand Protein Primary Structure • Calcium Coordination • 7 oxygen atoms from: • 3 mono-dentate side chains • 1 backbone carbonyl • 1 water H-bonded to side chain • 1 bi-dentate side chain • Specific geometry: pentagonal bi-pyrimid 7 5 9 3 1 12
S100 Proteins are Unique CaBPs • S100 proteins have a unique dimeric structure Potts. et al., 1995 2 slides
Dimer is the Basic Structural Unit • Interdigitated side chains • A single contiguous hydrophobic core 6 slides
S100 Protein Mechanism is Unique • S100 proteins have a unique dimeric structure • The mode of signal transduction must be distinct from calmodulin • Smaller changes in conformation • Target interactions are different Potts. et al., 1995
The Calmodulin Paradigm Ca2+ Target
Activation of Typical Ca2+ Sensors Accessible Hydrophobic Surface Calmodulin N-terminal Domain
S100 Response to Ca2+ Binding is Small • Significant change only in Helix III
Ca2+-induced Conformational ChangeS100s Are Different From Calmodulin Ca2+ Sensor S100B CaM-N • S100 protein response is much smaller than typical Ca2+ sensors
S100 Proteins Bind Helical Targets S100B/p53 • Shift in Helix III exposes binding site
MLCK S100s Must Bind Targets Differently p53 Calmodulin/MLC Kinase S100B/p53 • Calcium-dependent regulation of targets: kinases, kinase substrates, receptors (RAGE), fatty acids (arachidonic acid!)
CONCLUSIONS100 proteins are structurally(and functionally) different from classical Ca2+ sensors-BUT-They are very similar to each other!
All S100 Proteins HaveThe Same Basic Architecture S100A9 S100A8
Similarity Extends Beyond Architecture A6 B A7 A8 A12 Calbindin D9k!!
A Bonus: Accurate Homology Modeling S100A4 Ca2+-bound Ca2+-free • We now know the basic structure of every S100 protein!! So why don’t we know all of the answers?…it’s all about the details!
If they are so similar,what provides the functional specificity of S100 proteins?
Functional Specificity: CalmodulinDifferences in the Binding Sites for Different Proteins calmodulin caltractin • Extremely similar structures, but subtle details different
Functional Specificity: CalmodulinDifferences in the Binding Sites for Different Proteins opposite charge extra cleft extra pocket Caltractin Calmodulin • Differences in charge and shape of the binding surface
Functional Specificity: S100 ProteinsDifferent Binding Sites for Different Proteins S100A6 S100B • Differences in size and shape of the binding sites
Differences in Hydrophobic Surface Apo S100A6 Apo S100B Ca2+ loaded S100A6 Ca2+ loaded S100B • Differences in D hydrophobic surface induced by Ca2+ binding
Differences in Electrostatic Surface S100B-P53 S100A11-Annexin-I • Complemented by the properties of the target
Functional SpecificityDifferent Binding Modes for Different Proteins
Functional SpecificityDifferent Binding Modes for Different Proteins S100A10/annexin II S100A11/annexin I S100B/p53 S100A9/Chaps
Functional SpecificityA New Concept!! S100B-Ndr S100B-p53 • Different binding modes for the same • S100 protein with different targets!!
Functional SpecificityDiversity in Quaternary Structure? S100A9 S100A12 Dimer Tetramer
Functional SpecificityDiversity in Quaternary Structure? Hexamer Dimer
Functional SpecificityDiversity in Quaternary Structure? Tetramer Dimer Hexamer Octamer
Summary of Structural Concepts • The S100 architecture is unique • Mode of action differs from the CaM paradigm • Structures (apo, Ca2+-bound) are very similar • What provides functional specificity?
Factors Providing Functional Specificity • Sequence variability of residues at the surface alters the character of binding sites • The complementarity of the binding surface and target leads to different binding modes • Different modes for different proteins • Multiple modes for each protein? • Oligomerization may effect the presentation of binding surfaces to targets
Structures of Both Homodimers S100A8 S100A9 • C-terminal tail of S100A9 is missing • Only homology models of heterodimer
Analysis of S100A8/S100A9 Interactions with Cellular Targets
S100A8/A9-Arachidonic Acid Titration apo Ca2+ Ca2+ + AA • Ca2+-bound S100A8/S100A9 binds arachidonic acid
15N S100A9 15N S100A9 + AA R R 15N - Ca- CO - - -15N - Ca H H NMR Analysis of Binding of Arachidonic Acid by S100A8/A9
NMR Analysis of Binding of Arachidonic Acid by S100A8/A9 • Differential effects on protein signals • Discrete binding site for ligand • Still seeking to optimize
RAGEReceptor for Advanced Glycations Endproducts V C2 C1 Variable (V) and Constant (C) IGG domains Cell Membrane • EN-RAGE receptor ligand: an S100 protein!
Limited Proteolysis to Find Structural Domains of RAGE Intact S-RAGE Primary fragments • Three putative stable domains: V, C1 and C2
Protection in the Presence of Targets + S-RAGE • S-RAGE • The binding of S-RAGE stabilizes S100A8/A9
Proteolysis Immobilized S100 Wash Elute Mass Spec Identification Affinity Chromatography RAGE
S100/RAGE Affinity Chromatography * Not pure!! * S-RAGE RAGE domain A9 A8 St FT W1 W2 Elution • Retention of intact S-RAGE and stable domain
S100/RAGE Affinity Chromatography S-RAGE RAGE domains Pure Partial Full Affin.. Digestion • Purified protein gives same result!
Titration of S100A8/A9 With S-RAGE • Differential effects on protein signals • Discrete binding site for S-RAGE