Download

1 / 67
670 likes | 743 Views
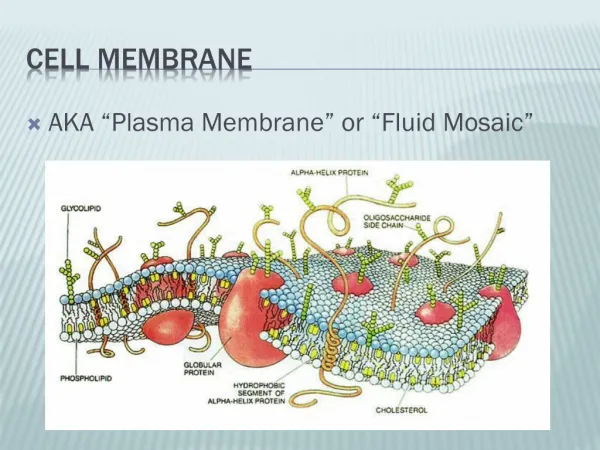
Control of Cell Volume and Membrane Potential. James Sneyd Auckland University, New Zealand. Basic reference: Keener and Sneyd, Mathematical Physiology (Springer, 1998). A nice cell picture. Basic problem. The cell is full of stuff. Proteins, ions, fats, etc.

E N D
Control of Cell Volume and Membrane Potential James Sneyd Auckland University, New Zealand Basic reference: Keener and Sneyd, Mathematical Physiology (Springer, 1998)
Basic problem • The cell is full of stuff. Proteins, ions, fats, etc. • Ordinarily, these would cause huge osmotic pressures, sucking water into the cell. • The cell membrane has no structural strength, and the cell would burst.
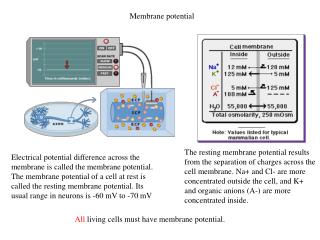
Basic solution • Cells carefully regulate their intracellular ionic concentrations, to ensure that no osmotic pressures arise • As a consequence, the major ions Na+, K+, Cl- and Ca2+ have different concentrations in the extracellular and intracellular environments. • And thus a voltage difference arises across the cell membrane. • Essentially two different kinds of cells: excitable and nonexcitable. • All cells have a resting membrane potential, but only excitable cells modulate it actively.
The cell at steady state • We need to model • pumps and exchangers • ionic currents • osmotic forces
Osmosis P1 P2 water + Solvent (conc. c) water At equilibrium: Note: equilibrium only. No information about the flow.
Na,K-ATPase The cell at steady state • We need to model • pumps and exchangers • ionic currents • osmotic forces Calcium ATPase I’ll talk about this a lot more in my next talk.
Active pumping • Clearly, the action of the pumps is crucial for the maintenance of ionic concentration differences • Many different kinds of pumps. Some use ATP as an energy source to pump against a gradient, others use a gradient of one ion to pump another ion against its gradient. • A huge proportion of all the energy intake of a human is devoted to the operation of the ionic pumps. • Not all that many pump models that I know of. It doesn't seem to be a popular modelling area. I have no idea why.
flux A Simple ATPase Note how the flux is driven by how far the concentrations are away from equilibrium
The cell at steady state • We need to model • pumps • ionic currents • osmotic forces
[S]i=[S’]i [S]e=[S’]e Vi Ve Permeable to S, not S’ The Nernst equation (The Nernst potential) Note: equilibrium only. Tells us nothing about the current. In addition, there is very little actual ion transfer from side to side. We'll discuss the multi-ion case later.
Only very little ion transfer spherical cell - radius 25 mm surface area - 8 x 10-5 cm2 total capacitance - 8 x 10-5 mF (membrance capacitance is about 1 mF/cm2) If the potential difference is -70 mV, this gives a total excess charge on the cell membrane of about 5 x 10-12 C. Since Faraday's constant, F, is 9.649 x 104 C/mole, this charge is equivalent to about 5 x 10-15 moles. But, the cell volume is about 65 x 10-9 litres, which, with an internal K+ concentration of 100 mM, gives about 6.5 x 10-9 moles of K+. So, the excess charge corresponds to about 1 millionth of the background K+ concentration.
Electrical circuit model of cell membrane How to model this is the crucial question
How to model Iionic • Many different possible models of Iionic • Constant field assumption gives the Goldman-Hodgkin-Katz model • The PNP equations can derive expressions from first principles (Eisenberg and others) • Barrier models, binding models, saturating models, etc etc. • Hodgkin and Huxley in their famous paper used a simple linear model • Ultimately, the best choice of model is determined by experimental measurements of the I-V curve.
Two common current models Linear model GHK model These are the two most common current models. Note how they both have the same reversal potential, as they must. (Crucial fact: In electrically excitable cells gNa (or PNa) are not constant, but are functions of voltage and time. More on this later.)
Electrodiffusion: deriving current models Poisson-Nernst-Planck equations. PNP equations. Poisson equation and electrodiffusion Boundary conditions
The short-channel limit If the channel is short, then L ~ 0 and so l ~ 0. This is the Goldman-Hodgkin-Katz equation. Note: a short channel implies independence of ion movement through the channel.
The long-channel limit If the channel is long, then 1/L ~ 0 and so 1/l ~ 0. This is the linear I-V curve. The independence principle is not satisfied, so no independent movement of ions through the channel. Not surprising in a long channel.
A Model of Volume Control Putting together the three components (pumps, currents and osmosis) gives.....
The Pump-Leak Model cell volume [Na]i pump rate Note how this is a really crappy pump model Na+ is pumped out. K+ is pumped in. So cells have low [Na+] and high [K+] inside. For now we ignore Ca2+ (horrors!). Cl- just equilibrates passively.
Charge and osmotic balance charge balance osmotic balance • The proteins (X) are negatively charged, with valence zx. • Both inside and outside are electrically neutral. • The same number of ions on each side. • 5 equations, 5 unknowns (internal ionic concentrations, voltage, and volume). Just solve.
Steady-state solution If the pump stops, the cell bursts, as expected. The minimal volume gives approximately the correct membrane potential. In a more complicated model, one would have to consider time dependence also. And the real story is far more complicated.
RVD and RVI Okada et al., J. Physiol. 532, 3, (2001)
Ion transport • How can epithelial cells transport ions (and water) while maintaining a constant cell volume? • Spatial separation of the leaks and the pumps is one option. • But intricate control mechanisms are needed also. • A fertile field for modelling. (Eg. A.Weinstein, Bull. Math. Biol. 54, 537, 1992.) The KJU model. Koefoed-Johnsen and Ussing (1958).
Steady state equations Note the different current and pump models electroneutrality osmotic balance
Transport control Simple manipulations show that a solution exists if Clearly, in order to handle the greatest range of mucosal to serosal concentrations, one would want to have the Na+ permeability a decreasing function of the mucosal concentration, and the K+ permeability an increasing function of the mucosal Na+ concentration. As it happens, cells do both these things. For instance, as the cell swells (due to higher internal Na+ concentration), stretch-activated K+ channels open, thus increasing the K+ conductance.
Inner medullary collecting duct cells IMCD cells Real men deal with real cells, of course. Note the large Na+ flux from left to right. A. Weinstein, Am. J. Physiol. 274 (Renal Physiol. 43): F841–F855, 1998.
Active modulation of the membrane potential: electrically excitable cells
Hodgkin, Huxley, and squid Hodgkin Huxley Don't believe people that tell you that this is a small squid
Resting potential • No ions are at equilibrium, so there are continual background currents. At steady-state, the net current is zero, not the individual currents. • The pumps must work continually to maintain these concentration differences and the cell integrity. • The resting membrane potential depends on the model used for the ionic currents. linear current model (long channel limit) GHK current model (short channel limit)
Simplifications • In some cells (electrically excitable cells), the membrane potential is a far more complicated beast. • To simplify modelling of these types of cells, it is simplest just to assume that the internal and external ionic concentrations are constant. • Justification: Firstly, it takes only small currents to get large voltage deflections, and thus only small numbers of ions cross the membrane. Secondly, the pumps work continuously to maintain steady concentrations inside the cell. • So, in these simpler models the pump rate never appears explicitly, and all ionic concentrations are treated as known and fixed.
Steady-state vs instantaneous I-V curves • The I-V curves of the previous slide applied to a single open channel • But in a population of channels, the total current is a function of the single-channel current, and the number of open channels. • When V changes, both the single-channel current changes, as well as the proportion of open channels. But the first change happens almost instantaneously, while the second change is a lot slower. I-V curve of single open channel Number of open channels
2a a S0 S1 S2 b 2b K+ channel gating S00 S01 S10 S11
2a a S00 S01 S02 S i j b 2b g d g d g d 2a a inactivation activation S10 S11 S12 b 2b Na+ channel gating activation inactivation
Experimental data: K+ conductance If voltage is stepped up and held fixed, gKincreases to a new steady level. four subunits rate of rise gives tn steady-state time constant Now just fit to the data steady state gives n∞
Experimental data: Na+ conductance If voltage is stepped up and held fixed, gNaincreases and then decreases. Four subunits. Three switch on. One switches off. steady-state time constant Fit to the data is a little more complicated now, but still easy in principle.
Hodgkin-Huxley equations applied current generic leak activation (increases with V) much smaller than the others inactivation (decreases with V)
An action potential • gNa increases quickly, but then inactivation kicks in and it decreases again. • gK increases more slowly, and only decreases once the voltage has decreased. • The Na+ current is autocatalytic. An increase in V increases m, which increases the Na+ current, which increases V, etc. • Hence, the threshold for action potential initiation is where the inward Na+ current exactly balances the outward K+ current.
Law of mass action Given a basic reaction k1 A + B C k-1 we assume that the rate of forward reaction is linearly proportional to the concentrations of A and B, and the back reaction is linearly proportional to the concentration of C.
Equilibrium Equilibrium is reached when the net rate of reaction is zero. Thus change in Gibb’s free energy or This equilibrium constant tells us the extent of the reaction, NOT its speed.
Enzymes • Enzymes are catalysts, that speed up the rate of a reaction, without changing the extent of the reaction. • They are (in general) large proteins and are highly specific, i.e., usually each enzyme speeds up only one single biochemical reaction. • They are highly regulated by a pile of things. Phosphorylation, calcium, ATP, their own products, etc, resulting in extremely complex webs of intracellular biochemical reactions.