Download

1 / 47
520 likes | 1.02k Views
Island Biogeography. Islands can serve almost as a laboratory for the study of biogeography. The biota of an island is simpler than that of a continental area, and the interactions are easier to understand. There are three types of islands:

E N D
Islands can serve almost as a laboratory for the study of biogeography. The biota of an island is simpler than that of a continental area, and the interactions are easier to understand.
There are three types of islands: • Islands that were originally part of a nearby continent, but were separated by rising sea levels (land-bridge islands). • Islands that are part of a volcanic island arc. • Seamount chains which formed over geological “hotspots”.
The types of islands have different characteristic flora and fauna. Islands formed by isolation from continents would have a biota which would be a subset of that on the continent. It would have changed, however, as the result of independent evolution and extinction. The biota of island arcs and hotspot island chains originally arrived by trans-ocean dispersal. In both cases, several islands exist at one time, creating the possibility for inter-island dispersal and a more complex pattern of evolutionary change.
Dispersal to islands is typically by a sweepstakes route,. The dispersing organisms share adapations that allow them to reach the island, rather than adaptations allowing them to live there once they reach it. This is one factor that restricts the diversity of life on islands.
Some flying animals, such as birds and bats, are capable of reaching even very distant islands.
Most land animals must rely on dispersal mechanisms like drifting on masses of debris. Although this process is likely rare, it certainly happens and has been documented for organisms like iguanas.
Long distance dispersal in plants is much more likely. A great many plants are adapted for such dispersal. In addition, the long distance dispersal of a plant species can typically be accomplished by a single spore or seed, where in animals it typically requires a pair of organisms or a pregnant female.
Some plants have developed seeds or fruits that can be carried in the sea without being harmed.
There is no doubt that the degree of isolation of an island or island group is a factor in determining the biota that it will support.
Jared Diamond showed that, on very remote islands, the number of species may be less than that predicted by equilibrium theory. This is because of the great difficulty in dispersing to these islands.
The ratio of observed species to the expected number declines with distance from New Guinea.
For conifers and flowering plants in the Pacific, diversity is much lower in the more isolated island groups of the central and eastern Pacific.
If we plot the number of genera vs. island area, it becomes clear that the two are related. The more isolated islands (represented by ) have fewer genera that less isolated islands of the same size.
Island life is probably more hazardous than that on the mainland. For one thing, catastrophic events have more severe effects. There is typically no place to hide.
Also, when a species is lost by extinction, it is more difficult to replace it be immigration than in a mainland situation. For these, and other reasons, islands tend to support fewer species than mainland areas of similar size.
Island populations are more likely to go extinct than those on mainlands, for several reasons: • Populations are typically smaller. • They have less genetic diversity. • They were not originally adapted to the island habitat.
Islands are typically depauperate in species richness relative to mainland areas of comparable size. Originally, this was explained by a nonequilibrium theory of island biogeography which stated that islands are depauperate because they have not had sufficient time to accumulate species by immigration.
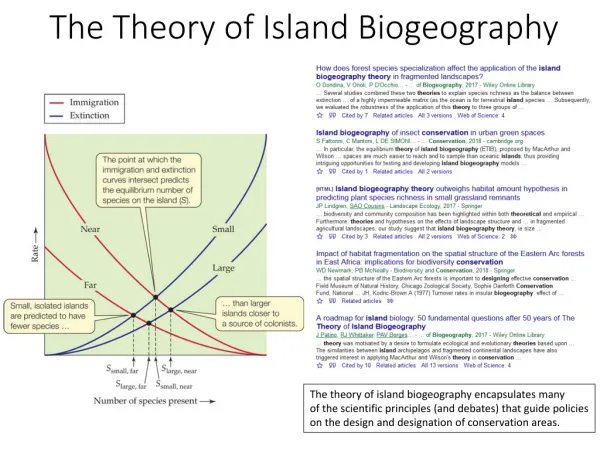
In 1963, Robert MacArthur and E.O. Wilson presented a new hypothesis to explain patterns of species richness on islands. Their equilibrium theory of island biogeography proposed that the lower number of species on islands was not the result of insufficient time, but rather the result of an equilibrium process peculiar to all islands.
The theory is based on the idea that, at any given time, the number of species on an island is the result of a balance between two processes: extinction and colonization.
When a new island forms, species begin to colonize. As more and more species accumulate, the colonization rate begins to decline. The extinction rate, on the other hand, begins to increase with increasing diversity.
At some point, the two processes balance each other, and the number of species on the island should stabilize. This equilibrium number is known as S
The equilibrium theory can also be used to explain the effect of size and distance on the number of species found on islands. Consider two islands of similar sizes but different distances from the mainland pool. Since extinction rates are a function of the available resources and should be related to the size of the island, we would expect them to be similar on the two islands. Colonization rates, however, should be greater for the island near the mainland than for the more distant island.
This should result in a difference in the equilibrium number of species, with Nnear > Nfar
A similar argument can be used to explain the effect of island size. If two islands are of relatively equal distance from the mainland, we can expect colonization rates to be similar. Extinction rates, however, should be greater on the smaller island. Therefore, we expect a higher equilibrium number of species on the large island.
So, the two approaches (nonequilibrium and equilibrium) make very different predictions about the nature of island species. • The equilibrium theory predicts that the number of species will not change over time. The nonequilibrium theory predicts that the number of species should increase with time. • The equilibrium species predicts that, although the number of species will remain relatively constant, the actual makeup of those species will change.
Several datasets have been developed that support the equilibrium theory. Jared Diamond looked at bird species on the Channel Islands off the California coast.
In 1969, E.O. Wilson and Daniel Simberloff conducted an experiment employing mangrove islets in the Florida Keys.
They surveyed a series of islands of differing sizes and distances from shore, concentrating on the arthropod fauna found on the islands.
Then, they defaunated the islands by enclosing them in plastic and pumping in methyl bromide to kill all the arthropods. They found that species increased for a while, then reached an asymptote approximately equal to the original number. But the makeup of the species had changed.
Following the publication of the theory, a number of other studies were conducted to examine its validity. A study on plant species on a group of islands off Britain showed that, in that case, the effect of size was indirect. Large islands had a greater degree of habitat heterogeneity, and therefore greater diversity.
Another factor is the nature of the islands. As mentioned earlier, some islands are of the land bridge type while others arose at sea and have never had a connection to the mainland. Oceanic islands confirm pretty closely to the patterns predicted by island biogeographic theory. Land bridge islands are a different story.
Land bridge islands begin with the species complement to be expected of a mainland area. Remember that this is typically more species than would be expected on an island of that size. So, over time, we expect the number of species to diminish. This is referred to as a relaxation fauna.
land-bridge island.. … or an oceanic island. So we see a different pattern for the number of species as a function of time for a:
Biogeographers and ecologists became anxious to apply these ideas to other island-like situations, particularly “habitat islands” on the mainland. A habitat island is a region of suitable habitat that is surrounded by uninhabitable area that serves as a barrier to dispersal.
The basin and range province in Nevada is an example of such a habitat island.
Other habitats that have been investigated using island biogeographic theory are vertebrates on mountaintops, invertebrates in caves, fish in lakes, and even herbivorous insects on plants. For the most part, the theory does not translate well. Plant diversity on mountaintops in the Cascade Range seems to increase with the age of the peak, rather than establish equilibrium.
There are some situations, however, where the theory does seem to apply. For example, there are species-area effects for birds and mammals inhabiting mountain ranges in the Basin and Range Province.
We can also see the effect of isolation. For mammals on montane habitat islands, increasing isolation (distance from the nearest habitat island) seems to lead to decreasing species richness. We also see evidence of a relaxation effect, resulting from the fact that these habitats were formerly more widespread.
Two important phenomena prevent habitat islands from conforming well to island biogeography theory: • Habitat diversity may be more important than area. This is true on oceanic islands as well, but the effect seems to be magnified in mainland habitat islands.
Bird species diversity in the Basin and Range is more closely correlated with habitat diversity than with area.
2. The formation of a habitat island in a mainland area creates an “edge effect”. This refers to a change in physical and biological characteristics of a habitat as one moves from the edge to the interior.
Island biogeography theory has had serious ramifications for conservation. One of the most significant anthropogenic influences on natural habitats has been the fragmentation of habitats into smaller and smaller “islands”. These fragmented habitats often show relaxation effects like some islands.
Mammal diversity in Mt. Rainier National Park has decreased by 26% between 1920 and 1976. This is probably due to the increasing fragmentation of forest habitats around the park due to logging.
We would like to use our increasing understanding of island biogeography theory to assist in solving practical conservation problems. Two areas are of particular interest: • Using the theory to predict the effect of anthropogenic habitat fragmentation. • Using our knowledge of species-area effects and relaxation faunas to design nature preserves that will maximize long-term species diversity.