Download

1 / 44
811 likes | 1.46k Views
Island Biogeography. Why study Islands?. First biologists and geographers studied them like Wallace (East Indies), Darwin (Galapagos Islands) and Hooker (Southern Ocean). Natural experimental plots which offer differences in sizes, number of species, isolation, number of predators.

E N D
Why study Islands? • First biologists and geographers studied them like Wallace (East Indies), Darwin (Galapagos Islands) and Hooker (Southern Ocean). • Natural experimental plots which offer differences in sizes, number of species, isolation, number of predators. • Interaction much less complex than in mainland habitats. • Due to their isolation evolutionary processes work at different rates • Little or no gene flow to dilute the effect of selection and mutation causing a very high level of endemism
Why study Islands? • Depending on scale and dispersal ability many habitats can be ‘Islands’ (lakes, mountaintops, etc.) • Islands can serve as natural field laboratories to study the relationship between area and species diversity • Part of unintentional experiments are habitat loss and introductions of invasive species by humans, often detrimental consequences • Only with a better understanding of species-area relationships can we design optimum conservation areas
What types of islands are there • Oceanic islands; which are located over oceanic plates and have never been connected to the continental shelf • Continental shelf islands: which are part of the continental shelf and can be connected to the mainland during periods of lower sea level • Habitat islands: distinct patches of terrestrial habitat surrounded by very different habitats but not water • Non-marine islands: which are somewhere between habitat and continental shelf islands in their level of isolation
Natural disturbances of islands • Any relative discrete event in time that removes organisms and opens up space which can be colonized by individuals of the same or a different species • Disturbances can be short term and frequently reoccurring like high winds or high rainfall • Some disturbances like ENSO events and hurricanes occurring every decade or more with larger impacts on islands • Other events occur only between 100 -1000 years for example volcanic eruptions, tsunamis or earthquakes
Implications of small founding populations • Typically the number of organisms arriving by a chance event on a remote island is small • Small founding populations containing only a subset of the source population’s biodiversity can cause a genetic bottleneck • Studies on Hawaiian fruit flies suggest that following the arrival of a single female with eggs on one of the islands, strong selection for females with less strict mate selection genes were more successful • Leading to a significant shift in gene frequencies allowing better adaptation to the new environment (Carson 2002)
Implications of small founding populations • The reduced genetic diversity in the founder population can also give rise to random genetic drift • Genetic drift by can lead to significant changes in a species genetic makeup even without further adaptation
Giants and dwarfs • The Galapagos and Indian Ocean tortoises were long regarded as typical island giants, but there have been large mainland species, only many are extinct due to humans • But a study on insular species of mammals found that 85% of island rodents are larger, possibly due to the absence of predators (Foster 1964, Arnold 1979)
Giants and dwarfs • On several islands in the Mediterranean dwarf hippopotami, elephants and deer existed several thousand years ago (Reyment 1983). • The record is the Maltan elephant which stood 1.5m shoulder height (Lister 1993) • The untested hypothesis is that on small islands there are less resources available for large herbivores and often no predators, therefore size reduction is an advantage • Maybe even human dwarf species Homo florensis on the Island of Flores (Brown et al 2004)
Giants and dwarfs • Three hypothesis for gigantism of island species (Schwaner & Sarre 1988) 1. Predation hypothesis: either there is selective release if no predation occurs or there is selective advantage to escape a window of vulnerability 2. Social-sexual hypothesis: due to high densities that occur among island populations, intraspecific competition among males and females selects for larger body size 3. Food availability hypothesis: increase in the mean and variance in food supply/demand ratio selects for giants
Loss of disperseability • An interesting aspect of many species which dispersed to islands is, that in many cases they lost their dispersal ability afterwards • Many birds became flightless, e.g. Aldabran rails, Dodo’s, Kakapo • Plants lost their ability of wind dispersal on near shore islands in BC (Cody and Overton 1996) and elsewhere • Flies lost their wings on Tristan da Cunha and Gough islands; elsewhere wing sizes are reduced • Original theory was this occurred due to preventing wind loss particular in insects, but Roff (1990,1994) found no clear relationship.
Ecological release on islands • Due to reduced competition or from other interacting organisms, like predators; leads to two main changes in newly arrived species • The loss of now unnecessary features (defensive traits, bold pattering, flight loss in many birds) • Examples are the Solomon Island rails which lost bold patterning and the ability to fly (Diamond 1991) • Many birds also reverted to simpler song patterns (Otte 1989) • Unfortunately many species also lost all fear of humans
Ecological release on islands • The second form of release is from close competitors, allowing the colonist to occupy not only different niches but also a wider array than its ancestral form (Cox & Ricklefs 1977) • It’s an important part for many scenarios of island evolution (e.g. adaptive radiation) • Examples are the Fijian fruit bats, that are more diurnal on islands without predatory eagles (Lomolino 1984) • Also the meadow vole is indiscriminate of habitat type on islands without predators (Lomolino 1984) • Nesting sites of several bird species on the Orkney Islands shifted from cliffs and trees to shrubs and flat ground
Adaptive radiation • Most well known examples are the Galapagos finches and the Hawaiian honey-creepers • The availability of empty niches is very important to adaptive radiation, allowing the diversification which sometimes leads to new species • There are also cases of non-adaptive radiation like the land snail genus Albinaria on the Island of Crete, which diversified without occupying different niches (Gittenberger 1991)
Island endemics • Many endemics to islands used to have a much wider distribution, but were replaced in other habitats, hence not all endemics have evolved in situ (palaeo-endemics) • One example is the St Helena Ebony; originates from a more widespread species 9 million years ago. Since then the family on the mainland has developed away from this species (Cronk 1987) • Whereas species evolved on islands are called neo-endemics • The issue: whether palaeo-endemics are more important for conservation due to a higher contribution to global biodiversity
Island endemics • The number of plant species endemic to the islands below (36,500) contribute 13.8% of the worlds higher plant species • About 7,000 of these are only found within a single island or island archipelago • The percentage of endemics are the highest for ancient continental islands like Madagascar and New Zealand • Islands contribute a disproportionate amount for their land area to global plant biodiversity
Island endemics • Land snails: only 8 archipelagos account between 7.7-9.0% of the world land snail species. In particular larger islands with higher elevation harbour many species (Groombridge 1992) • Insects: in Hawaii’ are alone about 1000 species of fruit flies (Wagner & Funk, 1995). • Lizards: Caribbean anoles are small arboreal insectivores and one of the larger and better studied vertebrate taxa. Out of 300 known Anolis species half occur on Caribbean islands (Losos 1994, 2004) • Birds: Galapagos finches and Hawaiian honeycreepers. 1750 species of birds are confined to islands, 17% of described species.
Species-isolation relationships • Another key factor determining the number of species on an island is the level of isolation • Islands of comparable sizes have a lower number of species if they are more isolated than habitat islands which are on continents (Wilson 1961)
Species-isolation relationships Williams (1981) found a decrease in the number of mainland bird species with increased distance from the mainland
Species-isolation relationships • Reasons for decline of species diversity with distance • Dependant on dispersal pathway, terrestrial mammals except bats can only disperse very limited distances (Lomolino, 1982)
Species-isolation relationships Bird species can disperse over larger distances, as seen in the example of resident land birds (Diamond 1972)
Species-isolation relationships • Dispersal abilities are also dependant on the type of reproduction a organism uses • Different estimates for ocean dispersal without human assistance is: freshwater fish 5km, elephants and other large mammals 50km, tortoises, snakes and rodents reached the Galapagos 1100km, bats and land birds reached Hawaii’ 3600km (Menard 1986) • Therefore the further an island is from the mainland the less species can disperse to it
Species-isolation relationships • Isolation from the mainland can also be changing over time • Example of lizard species on Islands in the Gulf of California (Wilcox 1978)
Species-area relationships • One of the most obvious traits of Islands are a limited number of species, more countable than on the mainland • The area available for species is also easier defined than on continents • Darlington (1957) found an empirical relationship between Island area and number of reptile and amphibian species in the West Indies
Species-area relationships • Darlington (1957) found an empirical relationship between Island area and number of reptile and amphibian species in the West Indies
Species-area relationships • As a log-log plot, it is not a curve but a straight line • As a rule of thumb with every 10 fold increase in size double the number of species are present. • S is number of Species • C is a constant which varies with the taxonomic group under study (taxa which consist of good dispersers (these species also typically have rapid population growth) will logically accumulate more species on an isolated island, all else being equal). • A is the area of the island, and the exponent z has been shown to be fairly constant for most island situations • Z represents a parameter for the slope of S and A on a log scale S = C A Z
Species-area relationships • Geographic variation in C has been observed and 'loosely' reflects the isolation of island groups typically studied • The presence or absence of major air or water circulation pathways nearby increases C • There are also effects of gross climatic difference, C is higher in the tropics than for islands at high arctic latitudes • C is also regarded as the the scaling factor
Species-area relationships • z in an all out treatment, is related to the distribution of abundances of species • Therefore the number of species expected if the total number of individuals increases, as it would on a larger island, and those species follow a Preston log-normal distribution of abundance (see May 1975) • Interpretation of these constant can be misleading (Lomolino 1989)
Species-area relationships • Many studies have looked at and compared z-values for different habitats • An early comparison (MacArthur and Wilson, 1967) found Islands to have z between 0.20-0.35 whereas non-isolated samples on continents or within large islands had a z of 0.12-0.17 • This suggests that any reduction in island area lowers the diversity more than a similar reduction of sample area in a contiguous mainland habitat • Other studies (Williamson 1988) have found a less clearly marked difference in z between mainland habitats and islands
Species-area relationships • Why might there be a difference in the species-area relationship between islands and isolated habitat areas on larger islands or continents? • The inclusion of transients in species counts from small 'islands‘ on continents • Species with large home ranges for example wolf with 400 square km, or even larger areas for seasonal migrants like caribou or large predatory birds • Such species might contribute to the number of species present but could not survive there if it would be a true island
Species-area relationships Species-area curves have been generated for a large variety of places and taxa, and the range of z values is remarkably small (Preston 1957, Williams 1953). Normally the relative abundance of species within a local biota fit log normal distribution
Species-abundance relationships • The curves indicate the presence of a few common species (the right hand end of the curve) and a larger number of species of intermediate abundance • The left hand end of the curve (the very rare species) are rarely included in studies, as they require a very high sampling effort
Species turnover • The Krakatau story and its lessons • A good record of recolonisation, particularly by bird species for the Krakatau Islands after the big volcanic eruption in 1883 • A rapid increase in bird species until 1920, after that number of species remained constant, but newcomers replaced already present species
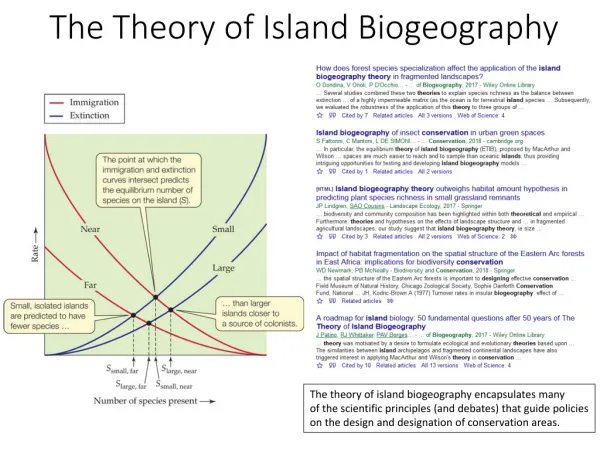
Equilibrium theory of island biogeography • Its based on the combination of species-area relationship, species-isolation relationship and species turnover (MacArthur and Wilson 1967). • It proposes that the number of species inhabiting an an island is based on the dynamic equilibrium between immigration and extinction. • The model is one of a dynamic equilibrium between immigration of new species onto islands and the extinction of species previously established.
Equilibrium theory of island biogeography • The formula is: St+1 = St+I+V-E • St is number of species at time t • I is the Immigration rate • V is additions through evolution • E is losses by extinction • The immigration rate is decreasing as there are fewer and fewer potential immigrant species remaining in the species pool P. This decrease is non-linear as the rate at which different species can disperse is different (e.g. tortoise vs bat) • The extinction rate increases non-linearly as factors like competition, predation, and parasitism become more important at higher species densities.
Tests of the equilibrium theory • In an experiment Simberloff (1976) censused terrestrial insect species on mangrove islands, and then cut the islands into smaller ones by creating 1m divides. This was sufficient to require jump dispersal from many insects • The smaller islands maintained a lower species number according with the equilibrium theory • Therefore in this study area as the only variable was a key determinant of number of species.
Is the world that simple? • Here are many criticisms of the ETIB • The theory ignores autoecology-but species are not exchangeable units (Armstrong 1982, Sauer 1969) • Data is rarely adequate for testing turnover (Lynch and Johnson 1974) • Most turnover involves transients (Simberloff 1976) • Turnover equilibrium has not been demonstrated (Gilbert 1980) • Immigration, extinction, and species pool are poorly defined (Williamson 1981,1989) • Ignores successional effects and pace, and the hierarchical links between taxa (Bush and Whitacker 1991)
Summary • Islands provide interesting study areas for the speciation, dispersal, colonization, evolution, radiation etc. • The simplified island world allows easier hypothesis testing than more connected continental habitats • Islands harbour a disproportional part of biodiversity
References • Main Sources • Whitaker RJ 1998. Island Biogeography, Ecology, Evolution, and Conservation, Oxford University Press. BOOK • Vitousek PM, Loope LL, Adsersen H (eds) 1995. Islands, biological diversity and ecosystem function. Springer. BOOK • Brown JH, Lomolino MV 1998 Biohepgraphy, second edition BOOK • Further reading • Carson HL Female choice in Drosophila: evidence from Hawaii and implications for evolutionary biology GENETICA 116 (2-3): 383-393 NOV 2002 • Foster JB 1964 Evolution of mammals on Islands, Nature, 202, 234-5 • Arnold EN 1979 Indian Ocean giant tortoises: their systematics and island adaptations. Philosophical Transactions of the Royal Society of London, series B, 286, 127-145 • Reyment RA 1983 Paleaontological aspects of island biogeography: colonization and evolution of mammals on Mediterranean islands. OIKOS, 41, 299-306 • Lister AM 1993 Mammoths in miniature. Nature, 362, 288-289 • Brown P, Sutikna T, Morwood MJ, Soejono RP, Jatmiko, Saptomo EW, Due RA A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia NATURE 431 (7012): 1055-1061 2004 • Schwaner TD, Sarre SD, 1988, Body size of Tiger Snakes in Southern Australia, with particular reference to Notechis ater serventyi (Elapidae) on Chappell Island. Journal of Herpatology, 22, 24-33 • Cody ML, Overton JMcC 1996 Shortterm evolution of reduced dispersal in island plant populations. Journal of Ecology, 84, 53-61 • ROFF DA THE EVOLUTION OF FLIGHTLESSNESS - IS HISTORY IMPORTANT EVOLUTIONARY ECOLOGY 8 (6): 639-657 1994 . • ROFF DA THE EVOLUTION OF FLIGHTLESSNESS IN INSECTS ECOLOGICAL MONOGRAPHS 60 (4): 389-421 1990 • Diamond JM 1991 A new species of rail from the Solomon islands and convergent evolution of insular flightlessness, The Auk, 108, 461-470 • Otte D, Endler JA (eds) 1989 Speciation and its consequences, Sinauer BOOK • Cox GW, Ricklefs RE, 1977 Species diversity, ecological release, and community structuring in Caribbean land bird faunas, Oikos, 28, 113-122
References • Lomolino MV 1984 Mammalian island biogeography: effects of area, isolation, and vagility. Oceologia, 61, 376-382 • Lomolino MV 1984 Immigrant selection, predation, and the distribution of Microtus pennsylvanicus and Blarina brevicauda on islands. The American Naturalist, 123, 468-483 • Gittenberger E 1991 What about non-adaptive radiation? Biological Journal of the Linnean Society, 43, 263-272 • Cronk, QCB 1989 The past and present vegetation of St. Helena. Journal of Biogeography, 16, 47-64 • Groombridge E (eds) 1992 Global biodiversity: status of the Earth’s living resources.BOOK • Wagner WL, Funk VA (eds) 1995 Hawaiian biogeography: evolution on a hot spot archipelago. Smithonian Press BOOK • Losos JB, Schoener TW, Spiller DA Predator-induced behaviour shifts and natural selection in field-experimental lizard populations NATURE 432 (7016): 505-508 NOV 25 2004 • LOSOS JB INTEGRATIVE APPROACHES TO EVOLUTIONARY ECOLOGY - ANOLIS LIZARDS AS MODEL SYSTEMS ANNUAL REVIEW OF ECOLOGY AND SYSTEMATICS 25: 467-493 1994 • Wilson EO 1961 The nature of the taxon cycle in the Melanesian and fauna. American Naturalist, 95, 169-193 • Williams MH 1981 Island populations BOOK • Lomolino MV 1993 Species-area and species-distance relationships of terrestrial mammals in the Thousand Island Region. Oecologia 54 72-75 • Diamond JM 1972 Biogeographic kinetics: Estimation of relaxation times for avifaunas of Southwest Pacific Islands. PNAS 69, 3199-3203 • Menard HW 1986 Islands BOOK • Wilcox BA 1978 Supersaturated island faunas: A species-age relationship for lizards on post-Pleistocene land-bridge islands. Science. 199, 996-998 • Darlington P.J.Jr 1957. Zoogeography: The Geographical distribution of animals. BOOK • May, R.M. 1975. Patterns of species abundance and diversity. in M.L. Cody and J.M. Diamond (eds.) Ecology and Evolution of Communities. Harvard Univ. Press, Cambridge, MA. pp.81-120.
References • Lomolino MV 1989 Interpretation and comparison of constants in the species-area relationship: An additional caution. American Naturalist 133, 71-75 • MacArthur RH, Wilson EO 1967 The theory of island biogeography. Monographs in population biology, no 1 BOOK • Williamson M 1989 The equilibrium theory today: True but trivial. Journal of Biogeography 16 3-4 • Preston FW 1957 Analysis of Maryland statewide bird counts. Maryland Birdlife 13, 63-65 • Williams CB 1953 The relative abundance of different species in a wild animal population. Journal of Animal Ecology 22, 14-31 • MacArthur RH Wilson EO 1963 An equilibrium theory of insular zoogeography. Evolution 17, 373-387 • Simberloff, D. 1976. Experimental zoogeography of islands: effects of island size. Ecology 57:629. • Armstrong P 1982 Rabbits (Oryctolagus cuniculus) on islands: a case study of successful colonization. Journal of Biogeography 9 353-362 • Sauer JD 1969 Oceanic islands and biogeographic theory: a review. The geographical review. 59, 582-593 • Lynch JD Johnson NV 1974 Turnover and equilibria in insular avifaunas, with special reference to the Califronian Channel Islands. The Condor, 76, 373-387 • Gilbert FS 1980 The equilibrium theory of island biogeography, fact or fiction? Journal of biogeography, 7, 209-235 • Bush MB Whittaker RJ 1991 Krakatau: colonization patterns and hierarchies, Journal of biogeography, 18 341-356