Download

1 / 67
690 likes | 1.22k Views
Neural Conduction. Ch. 4. Outline. Measuring the Membrane Potential The Ionic Basis of Resting Membrane Potential Four Factors Determine the Ionic Distribution that Underlies the Resting Membrane Potential Postsynaptic Potentials Generation of Action Potentials

E N D
Neural Conduction Ch. 4
Outline • Measuring the Membrane Potential • The Ionic Basis of Resting Membrane Potential • Four Factors Determine the Ionic Distribution that Underlies the Resting Membrane Potential • Postsynaptic Potentials • Generation of Action Potentials • Ionic Events Underlying Action Potentials • Conduction of Action Potentials • Changing Views on Dendritic Function
Measuring theMembrane Potential • to learn how information is sent from the dendrites and soma of a neuron to its terminals, researchers study a neuron’s membrane potential (the difference in electrical charge between the inside and outside of a neuron).
Measuring theMembrane Potential • to record a membrane potential two electrodes are needed: an intracellular electrode and an extracellular electrode • intracellular electrodes must be microelectrodes
The Ionic Basis of theResting Membrane Potential • when both intracellular and extracellular electrodes are outside a neuron, the difference between the electrical potentials at their tips is zero; when the intracellular electrode penetrates the neuron, the potential jumps to about -70 millivolts (the inside is 70 millivolts less than the outside)
The Ionic Basis of theResting Membrane Potential • The resting potential is thus, -70 millivolts • the resting potential exists because positively and negatively charged ions are distributed unequally on the two sides of the neural membrane: the concentration of Na+ and Cl- are higher outside the neuron, and the concentration of K+ and various negatively charged proteins are higher inside the neuron
Four factors determine the ionic distribution that underlies the resting potential
Ionic Distribution that Underlies the Resting Potential • four factors interact to produce the resting membrane potential; two passive (non-energy-consuming) factors act to distribute ions equally across the membrane (homogenizing factors) and one passive and one active factor to distribute ions unequally across the membrane
Random Motion (passive) • ions in solutions are in random motion • thus, any time that there is an accumulation of a particular class of ions in one area, the probability is increased that random motion will move ions out of this area (because there are more ions available to leave) and the probability is decreased that random motion will move more ions into the area (because there are fewer ions available to come in)
Electrostatic Pressure (passive) • like charges repel and opposite charges attract; therefore electrostatic pressure disperses any accumulation of positive or negative charges in an area
Differential Permeability of the Membrane (passive) • ions pass through membrane at special pores (made of proteins) called ion channels • when neurons are at rest, the membrane is: • totally resistant to the passage of protein ions, • extremely resistant to the passage of Na+ ions • and only slightly resistant to the passage of K+ ions and Cl- ions
Sodium-Potassium Pumps (active) • active (energy consuming) mechanisms in the neural membrane continuously transfer Na+ ions (that naturally leak in) out of the neuron and K+ ions (that leak outa) in • this occurs at a ratio of 3 Na+ moved out for every 2 K+ moved in; thus there is a loss of 1 positive charge every time the pump is activated
Post-synaptic Potentials • now that you understand the the state of resting neurons, let’s see how electrical signals are created in them
Post-synaptic Potentials • postsynaptic potentials are changes in the membrane potential produced by the action of neurotransmitters released by presynaptic neurons
Post-synaptic Potentials • Excitatory Postsynaptic Potentials (EPSPs) are depolarizations (decrease resting pot. by making more pos.); they increase the likelihood that a neuron will fire • Inhibitory Postsynaptic Potentials (IPSPs) are hyperpolarizations (increase rest. pot. by making more neg.); they decrease the likelihood that a neuron will fire
Post-synaptic Potentials • Postsynaptic potentials have three important properties: (1) They are graded (their amplitude is proportional to the intensity of the input; that is, stronger stimuli produce bigger EPSPs and IPSPs)
Post-synaptic Potentials (2) they are transmitted decrementally (as they passively spread from their site of generation, they get weaker as they go, like sound through air) (3) they are transmitted rapidly (like electricity through cable, so rapidly that transmission is regarded as being instantaneous)
Generation of Action Potentials • action potentials (neuron firing) are triggered at the axon hillock when a neuron is depolarized to the point that the membrane potential reaches about -65 mV; this is the threshold of excitation for many neurons
Generation of Action Potentials • Once threshold of activation of -65mV is reached, the Action Potential is reversal of the membrane potential from -70 mV to +50 mV in about one millisecond (Fig. 4.3)
Generation of Action Potentials • unlike EPSPs and IPSPs, APs are not graded; they are all-or-none (they occur full blown or not at all) • most neurons receive hundreds of synaptic contacts; what happens at any one synapse has very little effect on the firing of the neuron; whether or not a neuron fires is determined by the adding together (integration) of what goes on at many neurons
Generation of Action Potentials • There are two kinds of neural integration: • spatial summation:post synaptic potentials from multiple synapses produced almost synchronously • temporal summation: postsynaptic potentials produced in rapid succession from one synapse
Generation of Action Potentials • spatial summation: can involve EPSPs+EPSPs; IPSPs+IPSPs; or EPSPs+IPSPs • temporal summation: can involve EPSPs+EPSPs or IPSPs+IPSPs NOTE: EPSPs and IPSPs cannot temporally summate… due to the fact that just a single synapse is involved, and it must be either excitatory or inhibitory but not both
Generation of Action Potentials • in a functioning neuron, both spatial and temporal summation go on continuously; synapses closer to the axon hillock have a larger effect on firing due to decremental transmission of postsynaptic potentials
Ionic Events UnderlyingAction Potentials • What makes the membrane potential shoot from -70mV to +50 mV? (diagram in class)
Ionic Events UnderlyingAction Potentials • when the threshold of excitation (about -65mV) is reached, voltage-gated Na+ channels open momentarily, and Na+ ions rush into the neuron under tremendous pressure from both their concentration gradient and the electrostatic gradient; this drives the membrane potential to about +50mV
Ionic Events UnderlyingAction Potentials • At the same time, voltage-gated K+ channels slowly begin to open. Most of these channels open at about the time that the membrane potential is about +50mV. At this point, K+ ions are driven out by the +50mV charge and by their high internal concentration; this repolarizes the neuron and leaves it slightly hyperpolarized for a few milliseconds
Ionic Events UnderlyingAction Potentials • because only a few ions adjacent to the membrane are involved in the generation of an action potential, the resting potential is readily reestablished by the random motion of ions (and NOT the Na+/K+ pump!)
Ionic Events UnderlyingAction Potentials • for 1-2 milliseconds after an action potential is generated, another action potential cannot be generated no matter what kind of input that neuron receives; this is called the absolute refractory period. This is followed by the relative refractory period, during which time an action potential can only be elicited by high levels of stimulation.
Action Potentials are Nondecremental and Slow • once an AP is generated at the axon hillock, it is transmitted along the axon; the purpose of axons is to transmit APs from the soma to the terminal buttons of neuron
Action Potentials are Nondecremental and Slow • conduction of APs along an axon is not like the transmission of PSPs; conduction of EPSPs and IPSPs is passive (like electricity through a cable), thus it is instantaneous and decremental; whereas conduction of an AP along an axon is active, and therefore slower and nondecremental
Action Potentials are Nondecremental and Slow • when voltage-gated Na+ channels on the hillock membrane open, Na+ ions rush in and a full blown AP is generated
Action Potentials are Nondecremental and Slow • the electrical disturbance that is created is transmitted passively to the next Na+ channels along the axon (exposed in the Nodes of Ranvier); in response, the voltage-gated Na+ channels there open and another full-blown potential is generated
Action Potentials are Nondecremental and Slow • In reality, the sodium channels are so tightly packed that is is best to think if APs as waves of depolarization spreading down an axon; the AP is regenerated along the length of the axon
Action Potentials are Nondecremental and Slow • APs also spread from the hillock back through the cell body and dendrites, but because the ion channels in the cell body and dendrites are chemical-gated rather than voltage-gated, transmission of action potentials through cell bodies and dendrites is passive
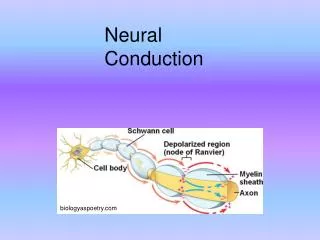
Myelin Increasesthe Speed of Conduction • many axons are myelinated by oligodendroglia in the CNS and by Schwann cells in the PNS; myelination insulates the semipermeable axon membrane, blocking the flow of ions through the axon at all but the Nodes of Ranvier; paradoxically this actually improves transmission
Myelin Increasesthe Speed of Conduction • In myelinated axons, APs travel passively (decrementally and rapidly) between the nodes of Ranvier; but at each node there is a pause while a full-blown AP is generated this is called a saltatory conduction
Myelin Increasesthe Speed of Conduction • because much of the transmission of APs in myelinated axons is passive (from node to node), transmission in myelinated axons is faster and it requires less energy • larger axons conduct faster; myelinated axons conduct faster (up to 60 meters per second in humans)
Changing Views onDendritic Function • dendritic function is more complex than was previously believed; for example, dendrites appear to be able to actively conduct action potentials, and many dendritic signals appear to be compartmentalized to just certain parts of the dendritic tree • the functional significance of these abilities remains to be determined; however, dendrites are clearly more than just passive conductors of action potentials
Synaptic Transmission Ch. 4 (cont’d)
Outline • Synaptic Contacts and Transmission • Neurotransmitters and receptors • Pharmacology of Synaptic Transmission
Structure of Synapses • In addition to axosomatic and axodendritic synapses, there are: (1) axoaxonic synapses (2) dendrodendritic synapses (3) dendroaxonic synapses (4) synapses between the main shafts of axons (5) nondirected synapses
Structure of Synapses • Some dendrodendritic synapses are reciprocal (they can transmit in either direction) • Many neurons have autoreceptors in their presynaptic membranes; these are stimulated by the neuron’s own neurotransmitter and are thought to mediate negative feedback • Some synapses occur on little buds on dendrites; these buds are called dendritic spines; other dendritic synapses occur right on the dendrite shaft
Structure of Synapses • Axoaxonic synapses mediate presynaptic inhibition; postsynaptic inhibition is mediated by axodendritic and axosomatic synapses
Structure of Synapses • Most synapses that are discussed in textbooks are directed synapses (synapses where the site of release and the target site are in close apposition) • There are also nondirected synapses; for example some presynaptic axons have a string-of-beads appearance and the neurotransmitter is widely dispersed from each bead to many targets in the general area
Synthesis, Packing and Transport of Neurotransmitter Molecules • There are two main types of neurotransmitters: • Small-molecule transmitters (synthesized in the cytoplasm of the terminal buttons and packed into vessicles by the Golgi complex) • Large-molecule (peptide) transmitters (synthesized in the soma by ribosomes and packed into vessicles by the soma’s Golgi complex and then moved down to the terminals by microtubules)
Synthesis, Packing and Transport of Neurotransmitter Molecules • There can be small and large molecule transmitters in the same terminal button; this is called coexistence
Release ofNeurotransmitter Molecules • The arrival of an AP at a terminal button opens voltage-gated calcium channels in the button membrane, and Ca++ ions enter the button
Release ofNeurotransmitter Molecules • The entry of the Ca++ ions causes the synaptic vessicles to fuse with the presynaptic membrane and empty their contents into synaptic cleft - a process called exocytosis