Download

1 / 51
510 likes | 710 Views
CHLOROPLASTS, CALVIN CYCLE, PHOTOSYNTHETIC ELECTRON TRANSFER AND PHOTOPHOSPHORYLATION (based on Chapter 19 and 20 of Stryer ). Photosynthesis. Photosynthesis. Light driven transfer of electron across a membrane Results in a proton gradient which drives the bonding of ADP and Pi to form ATP

E N D
CHLOROPLASTS, CALVIN CYCLE, PHOTOSYNTHETIC ELECTRON TRANSFER AND PHOTOPHOSPHORYLATION (based on Chapter 19 and 20 of Stryer )
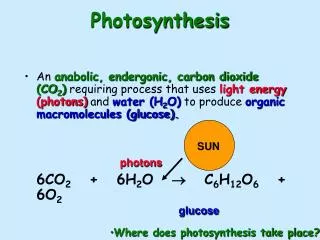
Photosynthesis Light driven transfer of electron across a membrane Results in a proton gradient which drives the bonding of ADP and Pi to form ATP Important in the production of Oxygen and the generation of carbon compounds that make aerobic metabolism possible Light CO2 + H2O (CH2O) + O2
Photosynthetic yield Huge amounts of free energy are stored annually amounting to more than 1010 tons of carbon in the form of carbohydrate “If a years yield were amassed in the form of sugar cane, it would form a heap over two miles high and with a base 43 square miles” - G.E.Fogge
STRUCTURE OF CHLOROPLASTS Typically 5um long. Surrounded by chloroplast envelope with outer and inner membrane and intermembrane space.
Inside is the stroma containing soluble enzymes, membranous structures called thylakoids, and a space inside the membranes called the lumen. Thylakoid membranes contain photosynthetic electron transfer chain and ATP synthase for photophosphorylation. They use light energy to synthesise NADPH and ATP (“light reactions”). The NADPH and ATP are used by the (Benson)-Calvin cycle to convert CO2 into sugar (“dark reactions”). The Calvin cycle is located in the stroma
PROKARYOTIC ORIGIN OF CHLOROPLASTS Phototrophic bacteria similar to cyanobacteria entered into an endosymbiotic association with the eukaryotic ancestors of algae and higher plants. Chloroplasts are no longer autonomous since most of their proteins are synthesised by nuclear genes.
Chlorophyll - photoreceptor Cyclic tetrapyrrole
CHLOROPHYLLS AND THE REACTION CENTRE The principal photoreceptor in chloroplasts is chlorophyll a bound to integral membrane proteins. Chlorophyll has two roles in trapping solar energy a) most chlorophylls absorb light energy and transfer it to a special pair of chlorophylls in a protein complex called a reaction centre. b) only the few chlorophylls forming the special pair mediate the transformation of absorbed light energy into chemical energy
Photosynthetic bacteria and photosynthetic reaction centres of green plants (PSI and PSII) have a common core Cytochrome C Prosthetic groups L M H
Electron chain in the photosynthetic bacterial reaction centre
Two photosystems generate a proton gradient and NADPH in oxygenic photosynthesis
PSII transfers electrons from water to plastoquinone and generates a proton gradient
Consists of three transmembrane protein complexes, two reaction centres (Photosystem II [PSII] and Photosystem I) and a cytochrome bf complex analogous to Complex III. These are linked by two mobile carriers a) plastoquinone (PQ) analogous to UQ links PSII and the cytochrome bf complex b) plastocyanin in the lumen is a small protein linking cytochrome bf complex and PSI (so analogous to cytochrome c )
The source of energy is light. This energy is absorbed and transferred to the two reaction centres, which use the energy to make an electron in the chlorophyll special pair more reducing. PS II takes electrons from H2O ( Eo 1/2 O2/H2O =+0.82V) and donates electrons to PQ(Eo PQ/PQH2 = 0V). Reaction centre has added 0.82V of reducing energy, using absorbed light energy to do so. Electrons then flow downhill in energy terms to plastocyanin (E0=+0.38V), where PSI uses light energy to take this electron and use it to reduce NADPH (Eo= -0.32V) via ferredoxin and ferredoxin-NADP+ oxidoreductase (FNR). Light causes electrons to flow from H2O to NADPH . This also leads to generation of a proton-motive force and thus ATP synthesis (photophosphorylation
There are three proton pumps 1) oxidation of water by PSII in the lumen releases protons 2) the cytochrome bf complex pumps protons from the stroma to the lumen 3) the reduction of NADP+ in the stroma removes protons So a proton-motive force is created with the lumen the P side, the stroma the N side. Protons flow back from the lumen to the stroma via an ATP synthase similar to that found on the IMM, leading to ATP synthesis.
Summary • Takes place in chloroplasts • Light absorption by chlorophyll Induces electron transfer • Two photosystems generate Proton Gradient and NADPH • Proton gradient drives ATP synthesis
BENSON-CALVIN CYCLE (The dark reaction) ATP and NADPH formed in the “light reactions” of photosynthesis are used to convert CO2 into hexose ( a 6 carbon sugar) and other carbon compounds. The “dark reactions” of photosynthesis start
6C 3C 4C 5C
1 The “dark reactions” of photosynthesis start with the reaction of CO2 and ribulose 1,5-bisphosphate (5C) to form two molecules of 3-phosphoglycerate (3C). This reaction is catalysed by ribulose 1,5-bisphosphate carboxylase/oxygenase (usually called rubisco). This carboxylase is also an oxygenase, because oxygen can also react with the enzyme to form phosphoglycolate (2C) and 3-phosphoglycerate. The recycling of phosphoglycolate leads to the wasteful loss of organic C as CO2 and the consumption of O2, in a process known as photorespiration.
ENERGY REQUIRED TO FIX CARBON IN THE CALVIN CYCLE Six rounds of the Calvin cycle are required to synthesise 1 hexose (6C) from 6 CO2. So a balanced equation for the net reaction of the Calvin cycle is 6 CO2 + 18 ATP + 12 NADPH + 12 H2O C6H12O6+ 18 ADP + 18 Pi + 12 NADP+ + 6H +
1,3-phosphoglycerate (3C) is converted into fructose 6-phosphate (6C) in four reactions which resemble similar steps in gluconeogenesis (the synthesis of glucose from non-carbohydrate sources such as lactate, amino acids and glycerol). Several of these reactions in the Calvin cycle and gluconeogenesis (synthesis of 6C) are shared with glycolysis (breakdown of 6C), except the essentially irreversible reactions in glycolysis have to be bypassed by new reactions. These steps in the Calvin cycle consume 2ATP and 2NADPH per 6C formed from 2(3C). The remaining task is to regenerate ribulose 1,5-bisphosphate (the CO2 acceptor), in other words to construct a 5C sugar from 6C and 3C sugars, a rearrangement that consumes 1ATP
STARCH AND SUCROSE ARE THE MAJOR CARBOHYDRATE STORES IN PLANTS Starch is a polymer of glucose residues. Starch is synthesized and stored in thechloroplasts. Sucrose is a dissacharide. Sucrose is synthesised in the cytosol, using triose phosphates (3C) exported from the chloroplasts.
THIOREDOXIN CO-ORDINATES “LIGHT” AND “DARK” REACTIONS Carbon dioxide assimilation and other biosynthetic reactions are switched on in the light by reduced thioredoxin A 12-kd protein reduced by ferredoxin which is reduced in turn by PSI in the light. The reduced thioredoxin reduces disulphide bridges which control the activities of biosynthetic enzymes.
Enzymes involved in carbohydrate degradation are switched off in the light when reduced thioredoxin reduces their disulphide bridges.The rate-limiting step in the Calvin cycle is that catalysed by rubisco. The activity of this enzyme increases in the light because the pH of the stroma increases from 7 to 8, and the level of Mg2+ increases in the stroma, as H+ are pumped into the lumen.