Download

1 / 49
490 likes | 530 Views
This article explores the process of protein synthesis and how the sequence of mRNA is translated into the sequence of a protein. It explains the genetic code and how it is translated from a "four-letter code" of mRNA into a "20-letter code" of proteins. The article also discusses the role of tRNAs as adapter molecules and the mechanisms of protein synthesis.

E N D
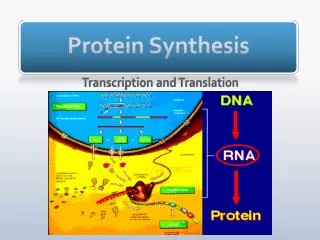
Translating the Message • How does the sequence of mRNA translate into the sequence of a protein? • What is the genetic code? • How do you translate the "four-letter code" of mRNA into the "20-letter code" of proteins? • And what are the mechanics like? There is no obvious chemical affinity between the purine and pyrimidine bases and the amino acids that make protein. • As a "way out" of this dilemma, Crick proposed "adapter molecules" - they are tRNAs!
The Collinearity of Gene and Protein Structures • Watson and Crick's structure for DNA, together with Sanger's demonstration that protein sequences were unique and specific, made it seem likely that DNA sequence specified protein sequence • Yanofsky provided better evidence in 1964: he showed that the relative distances between mutations in DNA were proportional to the distances between amino acid substitutions in E. coli tryptophan synthase
Elucidating the Genetic Code • How does DNA code for 20 different amino acids? • 2 letter code would allow for only 16 possible combinations. • 4 letter code would allow for 256 possible combinations. • 3 letter code would allow for 64 different combinations • Is the code overlapping? • Is the code punctuated?
The Nature of the Genetic Code • A group of three bases codes for one amino acid • The code is not overlapping • The base sequence is read from a fixed starting point, with no punctuation • The code is degenerate (in most cases, each amino acid can be designated by any of several triplets)
How the code was broken • Assignment of "codons" to their respective amino acids was achieved by in vitro biochemistry • Marshall Nirenberg and Heinrich Matthaei showed that poly-U produced polyphenylalanine in a cell-free solution from E. coli • Poly-A gave polylysine • Poly-C gave polyproline • Poly-G gave polyglycine • But what of others?
Getting at the Rest of the Code • Work with nucleotide copolymers (poly (A,C), etc.), revealed some of the codes • But Marshall Nirenberg and Philip Leder cracked the entire code in 1964 • They showed that trinucleotides bound to ribosomes could direct the binding of specific aminoacyl-tRNAs • By using C-14 labelled amino acids with all the possible trinucleotide codes, they elucidated all 64 correspondences in the code
Features of the Genetic Code • All the codons have meaning: 61 specify amino acids, and the other 3 are "nonsense" or "stop" codons • The code is unambiguous - only one amino acid is indicated by each of the 61 codons • The code is degenerate - except for Trp and Met, each amino acid is coded by two or more codons • First 2 codons of triplet are often enough to specify amino acid. Third position differs • Codons representing the same or similar amino acids are similar in sequence (Glu and Asp)
tRNAs • tRNAs are interpreters of the genetic code • Length = 73 – 95 bases • Have extensive 2o structure • Acceptor arm – position where amino acid attached • Anticodon – complementary to mRNA • Several covalently modified bases • Gray bases are conserved between tRNAs
Third-Base Degeneracy • Codon-anticodon pairing is the crucial feature of the "reading of the code" • But what accounts for "degeneracy": are there 61 different anticodons, or can you get by with fewer than 61, due to lack of specificity at the third position? • Crick's Wobble Hypothesis argues for the second possibility - the first base of the anticodon (which matches the 3rd base of the codon) is referred to as the "wobble position"
The Wobble Hypothesis • The first two bases of the codon make normal H-bond pairs with the 2nd and 3rd bases of the anticodon • At the remaining position, less stringent rules apply and non-canonical pairing may occur • The rules: first base U can recognize A or G, first base G can recognize U or C, and first base I can recognize U, C or A (I comes from deamination of A) • Advantage of wobble: dissociation of tRNA from mRNA is faster and protein synthesis too
AA Activation for Prot. Synth. • Codons are recognized by aminoacyl-tRNAs • Base pairing must allow the tRNA to bring its particular amino acid to the ribosome • But aminoacyl-tRNAs do something else: activate the amino acid for transfer to peptide • Aminoacyl-tRNA synthetases do the critical job - linking the right amino acid with "cognate" tRNA • Two levels of specificity - one in forming the aminoacyl adenylate and one in linking to tRNA
Aminoacyl-tRNA Synthetase Amino acid + tRNA + ATP aminoacyl-tRNA + AMP + PPi • Most species have at least 20 different aminoacyl-tRNA synthetases. • Typically one enzyme is able to recognize multiple anticodons coding for a single amino acids (I.e serine 6 different anticodons and only one synthetase) • Two step process: • Activation of amino acid to aminoacyladenylate • Formation of amino-acyl-tRNA
Specificity of Aminoacyl-tRNA Synthetases • Anticodon and structure features of acceptor arm of specific tRNAs are important in enzyme recognition • Synthetases are highly specific for substrates, but Ile-tRNA synthetase has 1% error rate. Sometimes incorporates Val. • Ile-tRNA has proof reading function. Has deacylase activity that "edits" and hydrolyzes misacylated aminoacyl-tRNAs
Translation • Slow rate of synthesis (18 amino acids per second) • In bacteria translation and transcription are coupled. As soon as 5’ end of mRNA is synthesized translation begins. • Situation in eukaryotes differs since transcription and translation occur in different cellular compartments.
Ribosomes • Protein biosynthetic machinery • Made of 2 subunits (bacterial 30S and 50S, Eukaryotes 40S and 60S) • Intact ribosome referred to as 70S ribosome in Prokaryotes and 80S ribosome in Eukaryotes • In bacteria, 20,000 ribosomes per cell, 20% of cell's mass. • Mass of ribosomes is roughly 2/3 RNA
Prokaryotic Ribosome Structure • E. coli ribosome is 25 nm diameter, 2520 kD in mass, and consists of two unequal subunits that dissociate at < 1mM Mg2+ • 30S subunit is 930 kD with 21 proteins and a 16S rRNA • 50S subunit is 1590 kD with 31 proteins and two rRNAs: 23S rRNA and 5S rRNA
Eukaryotic Ribosome Structure • Mitochondrial and chloroplast ribosomes are quite similar to prokaryotic ribosomes, reflecting their supposed prokaryotic origin • Cytoplasmic ribosomes are larger and more complex, but many of the structural and functional properties are similar • 40S subunit contains 30 proteins and 18S RNA. • 60S subunit contains 40 proteins and 3 rRNAs.
Ribosome Assembly • Assembly is coupled w/ transcription and pre-rRNA processing
Ribosome Structure • Crystal structure of ribosome is known • mRNA is associated with the 30S subunit • Two tRNA binding sites (P and A sites) are located in the cavity formed by the association of the 2 subunits. • The growing peptide chain threads through a “tunnel” that passes through the 40S (30S in bacteria) subunit.
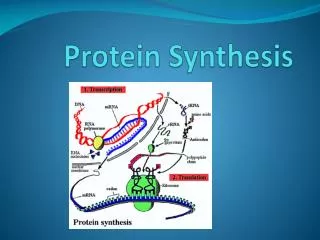
Mechanics of Protein Synthesis • All protein synthesis involves three phases: initiation, elongation, termination • Initiation involves binding of mRNA and initiator aminoacyl-tRNA to small subunit, followed by binding of large subunit • Elongation: synthesis of all peptide bonds - with tRNAs bound to acceptor (A) and peptidyl (P) sites. • Termination occurs when "stop codon" reached
Identification of Initiator Codon in Prokaryotes • Involves binding of initiator tRNA (N-formylmethionyl-tRNA) to initiator codon (first AUG) • The 30S subunit scans the mRNA for a specific sequence (Shine-Dalgarno Sequence) which is just upstream of the initiator codon. 16S RNA is involved in recognition of S-D sequence.
Prokaryotic Translational Initiation • Formation of Initiation complex involves protein initiation factors • IF-3 keeps ribosome subunits apart • IF-2 identifies and binds initiator tRNA. IF-2 must bind GTP to bind tRNA. • IF-1, IF-2, and IF-3 bind to 30S subunit to form initiation complex • Once 50S subunit binds initiation complex, GTP is hydrolyzed, initiator tRNA enters P-site and IFs disassociate
Eukaryotic Initiation of Translation • No S-D sequence. • CAP binding protein (CBP) 5’ end of mRNA by binding to 5’ CAP structure • An initiation complex forms with CBP, initiation factors and the 40S subunit. • The complex then scans the mRNA looking for the first AUG closest to the 5’ end of the mRNA • eIF-2 analogous to IF-2, transfers tRNA to P sight. GTP hydrolysis involed in release
Chain Elongation Three step process: • Position correct aminoacyl-tRNA at acceptor site • Formation of peptide bond between peptidyl-tRNA at P site with aminoacyl-tRNA at A site. • Shifting mRNA by one codon relative to ribosome.
Elongation Factor Tu (EF-Tu) binds to aminoacyl-tRNA and delivers it to the A site of the ribosome • When EF-Tu binds GTP a conformational change occurs allowing it to bind to aminoacyl-tRNA.
EF-Tu-tRNA complex enters the ribosome and positions new tRNA at A site. • If the anticodon matches the codon, GTP is hydrolyzed and EF-Tu releases the tRNA and then exits the ribosome.
Recycling of EF-Tu • After leaving the ribosome EF-Tu-GDP complex associates with EF-Tscausing GDP to disassociate. • When GTP bind to the EF-Tu/EF-Ts complex, EF-Ts disassociates and EF-Tu can bind another tRNA
Formation of Peptide Bond • Once the peptide bond forms, the mRNA band shifts to move the new peptidyl-tRNA into the P-site and moves the deaminacyl-tRNA from the E-site • Binding of EF-GTP to ribosome promotes the translocation • Hydrolysis of EF-GTP to EF-GDP is required to release EF from ribosome and new cycle of elongation could occur
More on elongation • Growing peptide chain then extends into the “tunnel” of the 50S subunit. • Floding of the native protein does not occur until the peptide exits the “tunnel” • Folding is facilitated by chaperones that are associated with the ribosome • To ensure the correct tRNA enters the A site, the 16S RNA is involved in determing correct codon/anticodon pairing at positions 1 and 2 of the codon.
Eukaryotic elongation process • Similar to what occurs in prokaryotes. • Analogous elongation factors. • EF-1a = EF-Tu docks tRNA in A-site • EF-1b = EF-Ts recycles EF-Tu • EF-2 = EF-G involved in translocation process
Peptide Chain Termination • Proteins known as "release factors" recognize the stop codon (UGA, UAG, or UAA) at the A site • In E. coli RF-1 recognizes UAA and UAG, RF-2 recognizes UAA and UGA. • RF-3 binds GTP and enhances activities of RF-1 and –2. • Presence of release factors with a nonsense codon at A site transforms the peptidyl transferase into a hydrolase, which cleaves the peptidyl chain from the tRNA carrier • Hydrolysis of GTP is required for disassociation of RFs, ribosome subunit and new peptide
Protein Synthesis is Expensive! • For each amino acid added to a polypeptide chain, 1 ATP and 3 GTPs are hydrolyzed. • This is the release of more energy than is needed to form a peptide bond. • Most of the energy is need to over-come entropy losses
Regulation of Gene Expression RNA Processing mRNA RNA Degradation 5’CAP AAAAAA Active enzyme Post-translational modification Protein Degradation
Regulation of Protein Synthesis Regulation could occur at two levels in translation • Initiation – formation of the initiation complex • Elongation – elongation could be stalled by if an mRNA contains “rare” codons
Regulation of Globin gene translation by heme • When heme is low, HCI kinase phosphorylates eIF-2-GDP complex, • GEF binds tightly to phosphorylated eiF-2-GDP complex • prevents recycling of eIF-2-GDP and stops translation
Regulation of the trp operon • Transcription and translation are tightly coupled in E. coli. • When Trp is aundant, transcription of the trp operon is repressed. • The mechanism of this repression is related to translation of the