Download
1 / 1
10 likes | 137 Views
Development of a sensitive force transducer for use with larval Dipteran muscle Julia Dettinger, Stephanie Albero, Jacob Krans Department of Biology, Mount Holyoke College, South Hadley, MA 01075. Physiology. Introduction. Electronics.

E N D
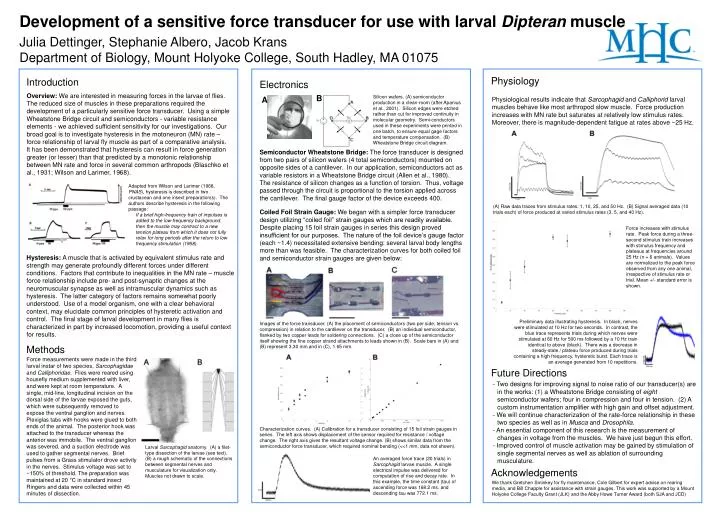
Development of a sensitive force transducer for use with larval Dipteran muscle Julia Dettinger, Stephanie Albero, Jacob Krans Department of Biology, Mount Holyoke College, South Hadley, MA 01075 Physiology Introduction Electronics Overview: We are interested in measuring forces in the larvae of flies. The reduced size of muscles in these preparations required the development of a particularly sensitive force transducer. Using a simple Wheatstone Bridge circuit and semiconductors - variable resistance elements - we achieved sufficient sensitivity for our investigations. Our broad goal is to investigate hysteresis in the motoneuron (MN) rate – force relationship of larval fly muscle as part of a comparative analysis. It has been demonstrated that hysteresis can result in force generation greater (or lesser) than that predicted by a monotonic relationship between MN rate and force in several common arthropods (Blaschko et al., 1931; Wilson and Larimer, 1968). B Silicon wafers. (A) semiconductor production in a clean-room (after Apanius et al., 2001). Silicon edges were etched rather than cut for improved continuity in molecular geometry. Semi-conductors used in these experiments were printed in one batch, to ensure equal gage factors and temperature compensation. (B) Wheatstone Bridge circuit diagram. A Physiological results indicate that Sarcophagid and Calliphorid larval muscles behave like most arthropod slow muscle. Force production increases with MN rate but saturates at relatively low stimulus rates. Moreover, there is magnitude-dependent fatigue at rates above ~25 Hz. Semiconductor Wheatstone Bridge: The force transducer is designed from two pairs of silicon wafers (4 total semiconductors) mounted on opposite sides of a cantilever. In our application, semiconductors act as variable resistors in a Wheatstone Bridge circuit (Allen et al., 1980). The resistance of silicon changes as a function of torsion. Thus, voltage passed through the circuit is proportional to the torsion applied across the cantilever. The final gauge factor of the device exceeds 400. Coiled Foil Strain Gauge: We began with a simpler force transducer design utilizing “coiled foil” strain gauges which are readily available. Despite placing 15 foil strain gauges in series this design proved insufficient for our purposes. The nature of the foil device’s gauge factor (each ~1.4) necessitated extensive bending: several larval body lengths more than was feasible. The characterization curves for both coiled foil and semiconductor strain gauges are given below: • Adapted from Wilson and Larimer (1968, PNAS), hysteresis is described in two crustacean and one insect preparation(s). The authors describe hysteresis in the following passage: • If a brief high-frequency train of impulses is added to the low-frequency background, then the muscle may contract to a new tension plateau from which it does not fully relax for long periods after the return to low frequency stimulation (1968). (A) Raw data traces from stimulus rates: 1, 10, 25, and 50 Hz. (B) Signal averaged data (10 trials each) of force produced at varied stimulus rates (3, 5, and 40 Hz). Force increases with stimulus rate. Peak force during a three-second stimulus train increases with stimulus frequency and plateaus at frequencies around 25 Hz (n = 6 animals). Values are normalized to the peak force observed from any one animal, irrespective of stimulus rate or trial. Mean +/- standard error is shown. Hysteresis: A muscle that is activated by equivalent stimulus rate and strength may generate profoundly different forces under different conditions. Factors that contribute to inequalities in the MN rate – muscle force relationship include pre- and post-synaptic changes at the neuromuscular synapse as well as intramuscular dynamics such as hysteresis. The latter category of factors remains somewhat poorly understood. Use of a model organism, one with a clear behavioral context, may elucidate common principles of hysteretic activation and control. The final stage of larval development in many flies is characterized in part by increased locomotion, providing a useful context for results. Force (Normalized) Preliminary data illustrating hysteresis. In black, nerves were stimulated at 10 Hz for two seconds. In contrast, the blue trace represents trials during which nerves were stimulated at 60 Hz for 500 ms followed by a 10 Hz train identical to above (black). There was a decrease in steady-state / plateau force produced during trials containing a high frequency, hysteretic burst. Each trace is an average generated from 10 repetitions. Images of the force transducer. (A) the placement of semiconductors (two per side; tension vs. compression) in relation to the cantilever on the transducer. (B) an individual semiconductor, flanked by two copper leads for soldering connections. (C) a close up of the semiconductor itself showing the fine copper strand attachments to leads shown in (B). Scale bars in (A) and (B) represent 3.30 mm and in (C), 1.65 mm. Methods Force measurements were made in the third larval instar of two species, Sarcophagidae and Calliphoridae. Flies were reared using housefly medium supplemented with liver, and were kept at room temperature. A single, mid-line, longitudinal incision on the dorsal side of the larvae exposed the guts, which were subsequently removed to expose the ventral ganglion and nerves. Plexiglas tabs with hooks were glued to both ends of the animal. The posterior hook was attached to the transducer whereas the anterior was immobile. The ventral ganglion was severed, and a suction electrode was used to gather segmental nerves. Brief pulses from a Grass stimulator drove activity in the nerves. Stimulus voltage was set to ~150% of threshold. The preparation was maintained at 20 °C in standard insect Ringers and data were collected within 45 minutes of dissection. Future Directions - Two designs for improving signal to noise ratio of our transducer(s) are in the works: (1) a Wheatstone Bridge consisting of eight semiconductor wafers; four in compression and four in tension. (2) A custom instrumentation amplifier with high gain and offset adjustment. - We will continue characterization of the rate-force relationship in these two species as well as in Musca and Drosophila. - An essential component of this research is the measurement of changes in voltage from the muscles. We have just begun this effort. - Improved control of muscle activation may be gained by stimulation of single segmental nerves as well as ablation of surrounding musculature. Characterization curves. (A) Calibration for a transducer consisting of 15 foil strain gauges in series. The left axis shows displacement of the sensor required for resistance / voltage change. The right axis gives the resultant voltage change. (B) shows similar data from the semiconductor force transducer, which required nominal bending (<<1 mm, data not shown). Larval Sarcophagid anatomy. (A) a filet-type dissection of the larvae (see text). (B) a rough schematic of the connections between segmental nerves and musculature for visualization only. Muscles not drawn to scale. An averaged force trace (20 trials) in Sarcophagid larvae muscle. A single electrical impulse was delivered for computation of rise and decay rate. In this example, the time constant (tau) of ascending force was 168.2 ms, and descending tau was 772.1 ms. Acknowledgements We thank Gretchen Sminkey for fly maintenance, Cole Gilbert for expert advise on rearing media, and Bill Chapple for assistance with strain gauges. This work was supported by a Mount Holyoke College Faculty Grant (JLK) and the Abby Howe Turner Award (both SJA and JCD)






















