Download

1 / 59
600 likes | 998 Views
Mekanisme Molekuler Induksi Tumor Crown Gall oleh Agrobacterium tumefaciens. IV. ANALISIS FUNGSIONAL PRODUK GEN AcvB. TIK : Mahasiswa mengetahui teknik-teknik analisa fungsional produk gen. Mahasiswa mampu mengolah hasil analisa fungsional produk gen. Waktu : 3 x(2x50 menit).

E N D
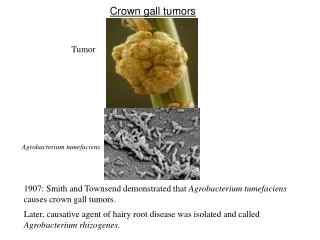
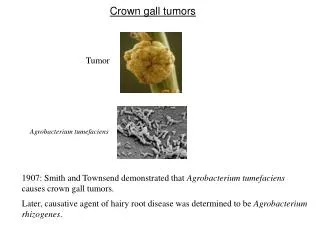
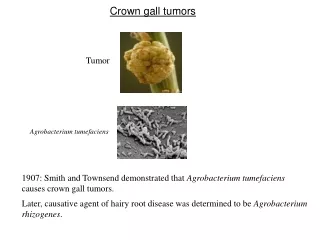
Mekanisme Molekuler Induksi Tumor Crown Gall oleh Agrobacterium tumefaciens
IV. ANALISIS FUNGSIONAL PRODUK GEN AcvB TIK : • Mahasiswa mengetahui teknik-teknik analisa fungsional produk gen. • Mahasiswa mampu mengolah hasil analisa fungsional produk gen. Waktu: 3x(2x50 menit)
Sub Pokok Bahasan: 4.1 Pendahuluan 4.2 Bahan dan Teknik Analisa Fungsional Produk Gen 4.2.1 Strain Bakteri dan Kondisi Kultur 4.2.2 Pemurnian Protein AcvB yang Diekspresi dalam E.coli. 4.2.3Pengujian Secara in vivo Transfer T-DNA dari A.tumefaciens ke Sel Kultur SuspensiTembakau 4.2.4Penyiapan Probe DNA 4.2.5Penyiapan Antisera 4.2.6 Pengujian Ratardasi Gel 4.2.7 Pembentukan Kompleks ssT-DNA-AcvB secara in vitro 4.2.8 Pendeteksian T strand kompleks yang mengandung AcvB dan protein VirE2 dalam periplasma A. Tumefaciens.
4.2.9 Isolasi Protoplas Daun Tomat 4.2.10 Penggabungan [32P]ssDNA ke dalam Protoplas Daun Tomat 4.2.11 Perlakuan terhadap Protoplast dengan Proteinase K atau DNase. 4.2.12 Pendeteksian Protein AcvB yang Tergabung dalam Protoplast dengan Menggunakan Teknik Imunofluorescence Secara Tidak Langsung 4.2.13 Perkiraan Stabilitas T-DNA:AcvB Kompleks dalam Protoplas Daun Tomat 4.2.14 Penentuan Distribusi Intraseluler dari [32P] T-DNA dalam Protoplas Daun Tomat 4.2.15 Pengujian Transfer T-DNA dari Strain A208 dan B119 ke Sel Tembakau (BY2) yang Mengekspresi Gen AcvB
4.3 Hasil Analisa Fungsional Produk Gen 4.3.1 Transfer T-DNA dari strain A208 dan B119 ke sel kultur suspensi tembakau 4.3.2 Interaksi protein AcvB dengan T-strand secara in vitro 4.3.3 Demonstrasi pembentukan T-DNA:AcvB kompleks secara in vitro dengan imunopresipitasi 4.3.4 Pendeteksian T-strand kompleks yang mengandung protein AcvB dan VirE2 dalam periplasma acetosyringone (AS) yang terinduksi A. Tumefaciens.
4.3.5 Stimulasi Penggabungan ssDNA ke dalam Protoplas Daun Tomat oleh Protein AcvB 4.3.6 Hambatan Penggabungan ssDNA ke dalam Protoplas Daun Tomat oleh serum anti-AcvB 4.3.7 T-DNA Tidak Hanya Terserap pada Permukaan Protoplas melainkan Tergabung ke Dalam Protoplas 4.3.8 Tujuan T-DNA:AcvB Kompleks dalam Protoplas Daun Tomat 4.3.9 Transfer T-DNA dari strain A208 dan B119 ke Tembakau (BY2) yang Mengekspresi Gen AcvB 4.4 Diskusi dan Kesimpulan
4.1 PENDAHULUAN • Gen acvB diekspresi secara berlebih pada Eschericia coli. • Sebuah protein 47 kDa diproduksi dan dilokalisasi dalam ruang periplasmik dari E. Coli. • Analisis susunan asam amino dari N-terminalnya menunjukkan bahwa peptida sinyal dari 24 asam amino dipotong dari protein pre-AcvB untuk menghasilkan protein 47 kDa dewasa/matang. • Hasil ini sesuai dengan penelitian bahwa susunan nukleotida gen acvB memiliki susunan pada terminal-5’ yang sepertinya merupakan sebuah susunan sinyal (gb. 9) (45, 88). • Susunan asam amino gen acvB menunjukkan sebuah protein yang dapat larut dan memiliki profil hidropati serta protein acvB yang diproduksi dalam E. Coli benar-benar dapat larut dalam air. • Anti serum untuk melawan protein acvB ditingkatkan dengan menggunakan seekor kelinci.
Analisis Western blot menggunakan anti serum mendeteksi adanya protein kDa 47 dalam periplasma strain A208 (acvB+) tapi tidak dalam strain B119 (acvB -) (gb. 20). • Gen acvB tidak terlibat dalam pengaturan gen vir pada Ti plasmid karena tidak terlihat perbedaan signifikan dalam induksi oleh acetosyringone dari gen fusi virB::lacZ, virD::lacZ, dan virE:: lacZ mengenai ada atau tidaknya gen acvB. Karena itu, gen acvB berbeda dari gen-gen virulen kromosom yang dilaporkan sebelumnya (chvD, chvE, miaA, ros, chvG dan chvI) yang terlibat dalam virulensi dengan cara mempengaruhi ekspresi gen vir dalam Ti plasmid. • T-strand (T-DNA dengan strand yang lebih rendah) terdeteksi pada strain B119 (acvB -) juga dalam strain A208 (acvB +) yang dikultur dalam medium induksi yang mengandung acetosyringone (penginduksi gen vir) (gb. 21). • Dalam bab 3 juga telah didemontrasikan bahwa gen acvB hanya ada dalam Agrobacterium, yang menunjukkan bahwa gen acvB terlibat dalam proses yang hanya berlangsung pada Agrobacterium tapi tidak pada bakteri lainnya.
Berdasarkan keseluruhan hasil-hasil percobaan tersebut di atas, ditunjukkan bahwa protein acvB terlokalisasi dalam ruang periplasmik dan terlibat dalam proses lebih lanjut setelah pembentukan T-strand selama induksi Crown gall. • Dalam bab ini, perlu dilakukan percobaan-percobaan untuk menentukan proses dimana protein acvB berfungsi, selain itu juga untuk mengetahui mekanisme fungsinya.
4.2 BAHAN DAN TEKNIK ANALISA FUNGSIONAL PRODUK GEN 4.2.1 Strain Bakteri dan Kondisi Kultur • Strain Agrobacterium tumefaciens dan Eschericia coli serta plasmid-plasmid yang digunakan dalam bab ini disusun dalam tabel 5. • A. tumefaciens ditumbuhkan baik dalam medium minimal AB (13) maupun dalam medium YEB pada suhu 28° C (75). • E. coli ditumbuhkan pada cawan agar L atau di dalam L-Broth pada suhu 37° C. • Untuk induksi A. tumefaciens, sel-sel hasil kultur semalaman pada medium YEB dikumpulkan dan selanjutnya disuspensi pada 5 x 108 sel/ml dalam medium AB-MES, pH 5.6 (27) dengan acetosyringone (100µM) dilanjutkan dengan inkubasi pada suhu 28° C selama 18 jam. • Konsentrasi antibiotik (µg/ml) yang digunakan dalam media adalah sebagai berikut: untuk A.tumefaciens; kanamisin, 100 dan untuk E. coli; ampisilin, 100 dan tetrasiklin, 20.
4.2.2 Pemurnian Protein AcvB yang Diekspresi dalam E. coli. • E. coli yang membawa plasmid pKE8 ditumbuhkan dalam 1 liter L Broth yang mengandung ampisilin pada suhu 37° C. • Setelah tumbuh selama 3 jam, IPTG ditambahkan ke dalam 1 mM konsentrasi akhir untuk menginduksi ekspresi gen acvB, dan kulturnya ditumbuhkan selama 12 jam lagi. • Sel-sel tersebut dipanen dengan sentrifugasi (7 k x g selama 10 menit), dibilas sekali dengan 50 ml NaCl 0.5%, disuspensi dalam 100 ml Tris-HCl dingin (ice cold) 20 mM, pH 8.0, yang mengandung sukrosa (20%) dan EDTA (2 mM). • Supensi tersebut diinkubasi selama 10 menit pada suhu 4° C dengan pengocokan kuat dan selanjutnya sel-sel tersebut dikumpulkan dengan sentrifugasi (7 k x g selama 10 menit). • Pelet tersebut disuspensi kembali sesegera mungkin dalam 200 ml air dingin (ice-cold) dan diinkubasi kembali selama 10 menit di bawah kondisi yang disebutkan tadi.
Suspensi sel disentrifugasi pada 7 x g selama 10 menit dan hasil supernatannya yang mengandung protein periplasmik dipisahkan dan dikonsentrasikan hingga 2ml dengan filtrasi melalui membran Diaflo YM-10 (Amicon). • Larutan terkonsentrasi tersebut diletakkan pada kolom DEAE Toyopearl (Tosoh) (1.2 x 10 cm) diekuilibrasi dengan Tris-HCl 50 mM, pH 7.5. Kolom tersebut dielusi dengan gradien linier dari 0 sampai NaCl 0.3 M dalam Tris-HCl 50 mM, pH 7.5, (50 ml). • Kandungan protein dalam fraksi-fraksi (1 ml) diukur pada absorbansi 280 nm. Sampel (15µl) dari fraksi-fraksi tersebut dianalisis dengan SDS-PAGE (10%) untuk mendeteksi adanya protein AcvB. • Fraksi yang mengandung protein AcvB dikumpulkan dan dikonsentrasi dengan filtrasi melalui membran Diaflo YM-10. Sampel yang terkonsentrasi diletakkan dalam kolom Ultrogel AcA44 (2.7 x 42 cm, IBF) diekuilibrasi dengan Tris-HCl, pH 7.5, dan dielusi dari kolom dengan larutan penyangga yang sama. • Fragmen yang mengandung protein AcvB dikumpulkan dan dikonsentrasikan dengan filtrasi.
4.2.3 Pengujian Secara in vivo Transfer T-DNA dari A.tumefaciens ke Sel Kultur Suspensi Tembakau • Ti plasmid biner, pIG121 (59), yang mengandung gen pelapor β-glucuronidase (GUS) dengan sebuah intron dalam susunan kode (coding sequence) di bagian hilir promoter 35 S virus mozaik kembang kol (35 S – Intron – GUS), dimasukkan ke dalam strains A208 dan B119 dengan perkawinan tiga induk (triparental mating) seperti yang dilaporkan (83). • Karena keberadaan intron, 35 S – Intron – GUS tidak mengekspresi aktivitas GUS yang dapat terdeksi dalam sel-sel A. tumefaciens, sementara 35 S – Intron – GUS mengekspresi aktivitas GUS dalam sel-sel tembakau (59). • Sel-sel strain A208 dan B119 hasil kultur semalaman yang mengandung pIG121 diinokulasi selama tiga hari ke dalam suspensi sel tembakau (BY2) yang tersubkultur.
Sel tembakau yang telah diinokulasi, diinkubasi dalam cawan petri tanpa pengocokan pada suhu 28° C selama 48 jam. • Selanjutnya, sel tembakau dicuci lima kali menggunakan medium LS (29) dengan cara sentrifugasi pada 78 x g selama 1 menit untuk menghilangkan sel bakterinya. • Sel tembakau yang yang telah dicuci disuspensi kembali dalam 12 ml medium LS. • Kemudian, aktivitas GUS sel tembakau diuji tingkatannya dengan menggunakan 6 ml suspensi tersebut menggunakan metode yang telah dilaporkan (38). • Aliquot (2.4 ml) dari suspensi yang tersisa disebarkan di permukaan cawan agar LS yang mengandung kanamisin (0.2 mg/ml) dan karbenisilin (0.5 mg/ml) dan diinkubasi dalam gelap pada suhu 25° C selama 6 minggu.
4.2.4 Penyiapan Probe DNA • Probe yang digunakan dalam pengujian retardasi gel disiapkan sebagai berikut. • Probe 1: T-DNA dari pTiT37 disonikasi dan digunakan untuk elektroforesis gel agarose (2%). Pita DNA pada 300-400 bp di permukaan gel dielusi secara elektronik dan diberi label dengan menggunakan alat pemberi label primer acak (Takara) dan menggunakan [32P] dCTP. Selanjutnya, DNA yang berlabel difraksinasi dengan elektroforesis gel poliakriamid (4%) dan DNA pada kisaran 50-200 bp dielusi dan digunakan sebagai probe 1 setelah didenaturasi dengan pemanasan. • Probe 2: fragmen DNA 357 bp dari wilayah promoter gen α-amilase padi (disediakan oleh Dr. S Mitsunaga di laboratorium kami) diberi label pada terminal 3’ dengan enzim Klenow dan [α-32P] ATP.
Probe 3: plasmid pUC18 diurai dengan HaeII dan fragmen 370 bp dipisahkan dengan menggunakan elektroforesis gel agarose (3%). Fragmen tersebut diberi label dengan alat pelabel terminal Megalabel (Takara) dan dengan [γ_32] ATP. Satu bagian dari DNA berlabel digunakan secara langsung sebagai probe DNA strand ganda, sementara bagian lainnya didenaturasi dengan pemanasan dan digunakan sebagai probe DNA strand tunggal. • Dalam percobaan imunopresipitasi, probe-probe DNA dipersiapkan sebagai berikut. • Probe 4, wilayah T-DNA Ti plasmid tipe nopalin (pTiC58) pertama-tama diurai dengan Sac II untuk menghasilkan fragmen 6 kb yang mana selanjutnya diurai dengan sal I menghasilkan fragmen 4 kb dan 2 kb. • Fragmen DNA 2 kb dimurnikan dengan menggunakan gel (gel-purified) dan diberi label menggunakan alat pelabelan primer acak (Takara) dan [α_32P] dCTP.
4.2.5 Penyiapan Antisera • Serum Anti-AcvB yang telah dipersiapkan sebelumnya digunakan dalam studi ini (45). • Serum Anti-VirE2 disediakan oleh Dr. Y. Machida (Universitas Nagoya).
4.2.6 Pengujian Retardasi Gel • Campuran penguji (10 µl) mengandung protein AcvB (14 µg) atau protein periplasmik (20-30µg) dalam Hepes-KOH 20 mM (pH 7.9), KCl 100 mM, MgCl2 12.5 mM, EDTA 0.2 mM, gliserol 20%, fenimetilsulfonil florida 1 mM, dithiothreitol 2 mM, 1 ng probe DNA berlabel [32P], dan 3µg poli(dI-dC). • Setelah inkubasi selama 30 menit pada suhu 20° C, campuran penguji diletakkan pada gel poliariamida (4%) yang disiapkan dengan menggunakan larutan yang berisi Tris-HCl 67 mM, pH 7.4, EDTA 10 mM, dan natrium asetat 33 mM yang telah diproses sebelumnya pada 20 mA selama 1 jam. • Elektroforesis dilakukan selama 2 jam pada 30 mA. Kemudian gel tersebut dikeringkan dan diautoradiografi. • Pita autoradiografi dilihat dengan menggunakan penganalisa gambar (BAS 2000, Fujix).
4.2.7 Pembentukan ssT-DNA: AcvB Kompleks Secara in vitro • ssT-DNA (1 µg, fragmen 2 kb Sal I dari T-DNA Ti plasmid tipe nopalin (pTiC58)) diinkubasi dengan protein AcvB (7 µg) dalam 22 µl larutan bufer pengikat (larutan penyangga Hepes-KOH 20 mM, pH 7.9, yang mengandung KCl (100 mM), MgCl2 (12.5 mM), EDTA (0.2 mM), gliserol (20%) fenilmetilsulfonil florida (1 mM) dan dithiothreitol (2 mM)) pada suhu 25° C selama 30 menit. • Selanjutnya, campuran reaksi diencerkan hingga 400 µl dengan larutan penyangga triton (Tris-HCl 50 mM, pH 0.8, NaCl 150 mM, MgCl2 1 mM, triton X-100 1% (v/v)) yang mengandung gliserol 10% (v/v) dan 15µl serum anti-AcvB atau serum preimun.
Campuran tersebut kemudian ditambahkan 40µl campuran protein A-Sepharose (125 mg/ml) (Farmasia) dalam larutan penyangga triton dan diinkubasi pada suhu 4° C selama 2 jam dengan pengadukan pelan. • Butiran protein A-Sepharose dipisahkan melalui sentrifugasi dan dibilas 3 kali dengan 1 ml larutan penyangga triton. • Kemudian butiran protein A-Sepharose diekstraksi dua kali dengan fenol kemudian dengan kloroform. • DNA dipisahkan dari ekstrak dengan pengendapan etanol dan diaplikasikan pada elektroforesis gel agarose. • DNA yang telah diurai kemudian dilewatkan pada membran nilon (Hybond N+, Amersham) di bawah kondisi tak terdenaturasi (75) dan dihibridisasi dengan probe 4 sebagaimana telah dilaporkan.
4.2.8 Pendeteksian T-strand Kompleks yang Mengandung Protein AcvB dan VirE2 dalam Periplasma A. tumefaciens. • Fraksi-fraksi protein periplasmik disiapkan dari sel A. tumefaciens baik yang terinduksi maupun tak terinduksi acetosyringone (As) dan kemudian diimunopresipitasi dengan serum anti-AcvB maupun serum preimun seperti yang telah dijelaskan di atas. • Kompleks imun itu dipergunakan dalam analisis blot Western dan Southern.
Untuk analisis blot Western, kompleks imun yang terikat dielusi dari butiran protein A-Sepharose dengan pemanasan pada suhu 95° C dalam sampel larutan penyangga SDS-PAGE selama 5 menit, diurai dengan menggunakan SDS-PAGE dan dikeringkan secara elektronik di atas sebuah membran PVDF ImmobilonTM (Milipori) pada 40 mA selama 1-2 jam. • Protein pada membran diwarnai secara imunologi (immuno-stained) menggunakan serum anti-AcvB maupun serum anti-VirE2 seperti yang dilaporkan (69). • Analisis blot Southern untuk mendeteksi T-strand dilakuksn dengan menggunakan probe 4 seperti metode yang dilaporkan (75).
4.2.9 Isolasi Protoplas Daun Tomat • Strain A208 memproduksi gall pada batang tanaman tomat. Karenanya, protoplas daun tomat digunakan dalam percobaan ini dimana penggabungan ssDNA ke dalam sel tanaman inang diteliti. • Daun tomat muda (1.0-1.5 g) dicuci dalam air distilasi, dipotong menjadi beberapa bagian dan diletakkan dalam 20 ml medium pengurai (pH 5.8) yang mengandung manitol (0.5 M, K-dextran sulfat (0.5%, w/v), driselase (0.5%, w/v, Kyowa Hatsuko) dan selulase ”Onozuka” RS (0.5%, w/v, Yakult). • Medium pengurai yang berisi daun tersebut disimpan dalam desikator kedap udara selama 2 menit untuk mempermudah jalannya infiltrasi medium pengurai ke dalam jaringan daun yang diikuti dengan inkubasi selama 2-3 jam pada suhu 25° C dan 50 osilasi/menit.
Jaringan daun yang telah terurai disaring melalui kasa nilon 80 µm dan filtratnya disentrifugasi untuk mendapatkan protoplas pada 46.4 x g selama 5 menit menggunakan rotor tipe swing-out. • Protoplas tersebut dibilas tiga kali dengan medium A (manitol 0.5 M dalam larutan penyangga K-fosfat-sitrat 25 mM, pH 5.5). • Terakhir, protoplas tersebut disuspensi kembali dalam medium A dan digunakan dalam percobaan selanjutnya.
4.2.10 Penggabungan [32P]ssDNA ke Dalam Protoplas Daun Tomat • [32P] ssDNA (probe 3, 1 ng) dipre-inkubasi dengan protein AcvB (7 µg) atau protein lainnya dalam larutan penyangga pengikat (binding buffer) selama 30 menit pada suhu 28° C dan dicampur dengan protoplas daun tomat (300µl, 4.0 x106 sel/ml) dilanjutkan dengan inkubasi pada suhu 28° C dalam waktu yang bervariasi dengan kocokan pelan. • Setelah inkubasi, protoplas dijadikan pelet pada 46.4 x g selama 5 menit dan dibilas tiga kali dengan medium A. Radioaktivitas dalam protoplasma yang diinkubasi ditentukan dengan sebuah scintillation counter untuk memantau penggabungan ssDNA ke dalam protoplas. • Untuk meneliti pengaruh dari serum anti-AcvB dalam penggabungan ssDNA, 15 µl baik serum anti-AcvB maupun serum preimun ditambahkan ke dalam campuran pre-inkubasi.
4.2.11 Perlakuan terhadap Protoplas dengan Proteinase K atau DNase. • Mula-mula protoplas ditambahkan dengan probe 4 kompleks dan protein AcvB dengan metode yang telah diuraikan di atas. • Kemudian, protoplas dicuci tiga kali dengan medium A. Protoplas yang telah dicuci disuspensi dalam 100 µl medium A (4.0 x 106 sel/ml) yang mengandung baik proteinase K (0.5 µg, Wako) atau DNAse (170 unit, Wako) dan diinkubasi pada suhu 28° C dengan waktu bervariasi dalam sebuah alat pengocok (50 osilasi/menit). • Protoplas yang telah diinkubasi dicuci satu kali dengan medium A dan kemudian radioaktivitasnya dihitung untuk mengikuti lepasnya T-DNA dari protoplas.
4.2.12 Pendeteksian Protein AcvB yang Tergabung dalam Protoplas dengan Menggunakan Teknik Imunofluorescence secara Tidak Langsung • Protoplas ditambahkan dengan ssT-DNA kompleks (2 kb, fragmen Sal I) dan protein AcvB seperti yang dijelaskan di atas. • Protoplas tersebut kemudian diinkubasi dalam larutan penyangga cacodylate-HCl 0.1 M, pH 7.2, mengandung glutaraldehid (3%, w/v) pada suhu 25° C selama 5 jam. • Kemudian protoplas dikumpulkan dengan sentrifugasi pada 3,000 x g selama 5 menit dan dicuci tiga kali dengan larutan penyangga SP (sorbitol 1.2 M dalam larutan penyangga K-fosfat, pH 6.5). • Kemudian, protoplas diinkubasi selama 15 menit pada suhu 25° C dalam larutan penyangga SP yang mengandung susu skim (5%, w/v) dan polioksietilen sorbitan monolaurat (Twee-20) (0.05%, w/v) untuk mengurangi ikatan yang tidak spesifik. • Protoplas yang telah diinkubasi dipisahkan dengan sentrifugasi, dibilas dan disuspensi kembali dalam larutan penyangga SP. Tahap-tahap selanjutnya dilakukan dalam ruangan lembab pada suhu 25° sesuai metode yang dilaporkan (30).
Dua puluh mikroliter suspensi protoplas (8 x 104 sel) dilekatkan di permukaan cawan kaca dengan poli (L-lisin) (0.1%, Sigma) selama 30 menit dan kemudian diinkubasi baik dengan serum anti-AcvB maupun serum preimun (1:500) dalam 20 µl larutan penyangga PBS (Na2HPO4, NaH2PO4 13 mM, NaCl 75 mM) yang mengandung B.S.A. (1%), triton X-100 (0.1%), dan NaN3 (0.1%), (larutan penyangga PBT) dan diinkubasi selama 1.5 menit dengan flourescein isotiosianat (FITC) coupled goat anti-rabbit IgG (1:500) (Sigma). • Protoplas dicuci tiga kali dengan larutan penyangga PBS, dipenuhi dengan larutan Dabco (diazabisiklooktana) (0.233 g Dabco, 800 ml air, 200 ml Tris-HCl 1 M, pH 8.0, dan gliserol 9 ml), ditutup dengan kaca penutup dan diamati di bawah mikroskop laser confocal (Olympus LSM-GB 200), dengan filter BP 535, O515 dan UV365 serta lensa LSM Uvpo40.
4.2.13 Perkiraan Stabilitas T-DNA:AcvB Kompleks dalam Protoplas Daun Tomat • [32P] T-DNA (1.0 x 106 cpm) diinkubasi bersama protein AcvB pada suhu 25° C selama 30 menit sebagaimana digambarkan di atas. • Campuran reaksi yang telah diinkubasi dibagi menjadi dua aliquot. Satu aliquot digunakan untuk elektroforesis gel poliakrilamid (4%). • Autoradiogram disiapkan dari gel tersebut dan dianalisa dengan menggunakan alat penganalisa gambar (Bas 2000-Fujix) untuk menentukan jumlah T-DNA:AcvB kompleks dan T-DNA bebas dalam campuran reaksi yang telah diinkubasi tersebut. • Enam puluh satu persen dari T-DNA berada dalam bentuk kompleks dan 39% dalam bentuk bebas. Sisa aliquot (72.5 µl, 5.0 x 105 cpm) dari campuran reaksi yang telah diinkubasi, diimunopresipitasi dengan menggunakan serum anti-AcvB.
Ikatan radioaktivitas dengan butiran protein A-Sepharose ditentukan setelah tiga kali pencucian; yaitu 1.7 x 104 cpm, 3.4% dari total radioaktivitas dalam aliquot. Dari kedua angka di atas, efisiensi imunopresipitasi dari T-DNA:AcvB kompleks dengan serum anti-AcvB dihitung sebesar 5.6%. • [32P] T-DNA (probe 4) dipre-inkubasi bersama protein AcvB dan digabungkan ke dalam protoplas di bawah kondisi yang sama dengan yang telah dipaparkan di atas. • Selanjutnya, protoplas tersebut diinkubasi pada suhu 25° C selama 1 jam kemudian dicuci sebanyak tiga kali. • Protoplas yang telah dicuci dikacaukan dengan menggunakan homogenizer teflon dan disentrifugasi pada 19 k x g selama 10 menit. Radioaktivitas dalam fraksi supernatan (sitosol) dihitung. • Aliquot fraksi sitosol diimunopresipitasi dengan serum anti-AcvB dan ikatan radioaktivitasnya pada butir protein A-Sepharose dihitung. Jumlah T-DNA:AcvB kompleks dalam fraksi sitosol dihitung dengan menggunakan perhitungan/ jumlah yang sudah didapat dan efisiensi imunopresipitasi (5.6%).
4.2.14 Penentuan Distribusi Intraseluler dari [32P] T-DNA dalam Protoplas Daun Tomat • Protoplas ditambahkan dengan T-DNA:AcvB kompleks dan diinkubasi pada suhu 25° C dalam rentang waktu yang bervariasi dalam medium Murashige-Skoog bersuplemen sukrosa (3%, w/v), inositol (100 µg/ml), asam nikotinik (0.5 µg/ml), tiamin-HCl (0.1 µg/ml) dan kinetin (3 µg/ml). • Protoplas yang telah diinkubasi dicuci sebanyak satu kali dengan medium A. Protoplas yang telah dicuci dikacaukan dan dipecah menjadi fraksi-fraksi inti (nuclear) (fraksi pelet) dan fraksi supernatan sesuai metode yang telah dilaporkan dengan sedikit perubahan (28).
Protoplas (1.3 x 106 sel) disuspensi dalam 1.5 ml piperazin-N 10 mM, larutan penyangga N’-bis (asam etanesulfonik) (PIPES)-KOH, pH 7.0, mengandung 2-metil-2,4-pentanediol (heksilen glikol, 1 M), MgCl (10 mM), triton x-100 (0.5%, v/v), 2-mercaptoetanol (5 mM) dan fenil-metilsulfonil fluoride (0.8 mM) dan dikacaukan dengan cara melewatkannya melalui dua lapisan nilon berpori 80 dan 20 µm. • Protoplas yang telah dikacaukan, disentrifugasi pada 3,000 x g selama 10 menit untuk mendapatkan supernatan dan fraksi endapannya.
Fraksi endapan dicuci sekali dengan larutan penyangga pencuci nuklei (nucleiwashing buffer) (larutan penyangga PIPES-KOH 10 mM, pH 7.0, yang mengandung heksilen glikol (0.5 M), MgCl2 (10 mM), 2-mercaptoetanol (5mM) dan fenilmetilsulfonil fluoride (0.8 mM) bersuplemen triton X-100 (0.5%, v/v) dengan cara sentrifugasi pada 3,000 x g selama 5 menit dan sekali dengan larutan penyangga nuklei. • Fraksi endapan yang telah dicuci digunakan sebagai fraksi inti (nuclear) dalam studi ini. • Untuk meneliti kemurnian fraksi inti, fraksi tersebut diuji kandungan katalase (penanda peroksisom), fumarase (penanda mitokondria), klorofil (penanda kloroplas), kandungan DNA dan protein dengan metode yang telah dilaporkan sebelumnya; yaitu katalase (50), fumarase (31), klorofil (41), DNA (25), dan protein (6). Radioaktivitas dalam fraksi inti (endapan) dan supernatan dari protoplas yang telah diinkubasi dalam rentang waktu berbeda ditentukan untuk mengetahui perubahan waktu dalam distribusi intraselular [32P] T-DNA
4.2.15 Pengujian Transfer T-DNA dari Strain A208 dan B119 ke Sel Tembakau (BY2) yang Mengekspresi Gen AcvB • Plasmid biner yang mengandung gen acvB tersusun seperti dalam gambar 22. • Sebuah fragmen DNA 1931 bp yang mengandung gen acvB yang diklon dalam pB140Δ4 diiurai dengan KpnI dan dipotong kasar (blunt ended) dengan menggunakan T4 DNA polimerase (Takara). • Setelah itu, fragmen DNA yang mengandung gen acvB dipotong dengan penguraian menggunakan SacI. Fragmen hasilnya disisipkan ke dalam daerah SmaI-SacI (Sst) dari vektor plasmid biner, pBI121. • Dalam susunan ini, gen acvB ditempatkan tepat di hilir promoter cAMV35S dan menggantikan gen β-glucuronidase (GUS) yang hilang.
Plasmid yang dihasilkan, pMW24, dimasukkan dalam E. Coli JM109 dengan transformasi dan kemudian dipindahkan ke dalam A. tumefaciens (LBA4404 yang mengandung plasmid pAL4404) dengan triparental mating (83). • Strain yang dihasilkan, LBA4404 (pAL4404, pMW24) digunakan untuk transformasi sel tembakau (BY2) seperti yang telah disebutkan di atas. Ekspresi gen acvB dalam sel BY2 dikonfirmasi dengan analisis blot Western dengan menggunakan serum anti-AcvB. • Sel tembakau yang mengekspresi gen acvB diinokulasi dengan A208 (pIG121) atau B119 (pIG121) menurut prosedur yang telah dipaparkan di atas. • Aktivitas GUS sementara (transient) dalam sel tembakau yang telah ditransformasi diuji kadarnya dengan metode yang telah dilaporkan (38).
Untuk menegasan lebih lanjut aktivitas GUS dalam sel tembakau yang bertransformasi, pengujian histokemikal untuk ekspresi GUS juga dilaksanakan menurut prosedur yang telah dilaporkan (16) dengan sedikit perubahan. • Sel tembakau yang telah bertransformasi (BY2) dicuci lima kali pada medium LS dengan sentrifugasi pada 78 x g selama 1 menit untuk menghilangkan sel-sel bakteri. Suspensi sel tembakau yang sudah dicuci tersebut dikumpulkan menggunakan sentrifugasi pada 78 x g selama 1 menit dan kemudian disuspensi kembali dalam 300 µl larutan penyangga yang mengandung K3[Fe(CN)6] 0.5 mM, K4[Fe(CN)6.3H2O] 0.5 mM, 5-bromo-4-kloro-3-indolil glukuronidase (X-gluc) 1.9 mM dan 0.3% (v/v) Triton X-100. • Suspensi sel diinkubasi pada suhu 37° C selama 12 jam. Sel yang mengekspresi GUS tampak sebagai sel yang berwarna biru.
4.3 HASIL ANALISIS FUNGSIONAL PRODUK GEN 4.3.1 Transfer T-DNA dari Strains A208 dan B119 ke Sel Kultur Suspensi Tembakau • Ti plasmid biner, pIG121 (59), yang mengandung gen reporter β-glukuronidase (GUS) dengan sebuah intron di dalam susunan kodenya begitu pula gen yang resisten terhadap kanamisin, dimasukkan ke dalam strain A208 dan B119 dan diinokulasi ke dalam sel kultur suspensi tembakau (BY2). • Setelah inkubasi selama 48 jam, kultur tersebut dibebaskan dari sel A. tumefaciens dan diuji aktivitas GUS-nya. • Ekspresi GUS sementara (transient) yang tinggi (41.9 pmoles MU/menit/mg protein) terdeteksi pada sel yang diinokulasi dengan strain A208 (pIG121), sedangkan ekspresi GUS yang sangat kecil (3.9 pmoles MU/menit/mg protein) terlacak pada sel yang diinokulasi dengan strain B119 (pIG121). • Hasil ini menunjukkan bahwa transfer DNA ke dalam sel tanaman inang terganggu oleh strain B119.
Aliquot sisa dari suspensi sel yang telah terinokulasi dikultur pada cawan agar medium LS yang mengandung kanamisin (0.2 mg/ml) dan karbenisilin (0.5 mg/ml) selama 6 minggu. • Pertumbuhan sel yang baik teramati pada sel tembakau yang diinokulasi oleh strain A208 (pIG121), sedangkan tidak ada pertumbuhan sel yang ditemukan pada sel tembakau yang diinokulasi dengan strain B119 (pIG121) (gb. 23). • Hasil ini menunjukkan bahwa transfer T-DNA dan integrasinya ke dalam genom terjadi pada sel yang diinokulasi dengan strain A208 (pIG121), tetapi tidak terjadi pada sel tembakau yang diinokulasi dengan strain B119 (pIG121), hal ini menegaskan hasil pengujian aktivitas GUS yang telah dipaparkan sebelumnya.
4.3.2 Interaksi Protein AcvB dengan T-strand secara in vitro • Untuk meneliti apakah protein AcvB dapat terikat pada T-strand, dilakukan pengujian ratardasi gel (gel retardation assay) (gb. 24). • Aktivitas pengikatan pada ssT-DNA strand tunggal dideteksi dengan fraksi protein periplasmik dari E.Coli yang mengandung plasmid pKE8 (lane 4, pada panel A). • Disisi lain, tidak terdeteksi adanya aktivitas pengikatan T-DNA dengan fraksi protein periplasmik pada E. coli yang mengandung plasmid vektor, pKE233-3 (lane 3 pada panel A) dan BSA atau bovine serum albumin (lane 2 pada panel A).
Hasil tersebut menunjukkan bahwa protein AcvB memiliki aktivitas pengikatan ssT-DNA. Bahkan, protein AcvB yang dimurnikan menunjukkan adanya aktivitas pengikatan ssT-DNA (lane 2 pada panel B). • Protein AcvB juga terikat pada fragmen DNA pada daerah promoter dari gen α-amilase padi (lane 1, pada panel C). • Hasil-hasil ini mengindikasikan bahwa protein AcvB tidak menunjukkan susunan yang spesifik dalam aktivitas pengikatannya. Percobaan yang serupa dengan menggunakan DNA strand ganda (ds) menunjukkan tidak adanya aktivitas pengikatan (lane 3, pada panel D).
4.3.3 Demonstrasi Pembentukan T-DNA:AcvB Kompleks secara in vitro dengan Imunopresipitasi • Protein AcvB menunjukkan adanya aktivitas pengikatan ssDNA dalam pengujian ratardasi gel (gel retardation assay) sebagaimana disampaikan sebelumnya. • Untuk menegaskan lebih lanjut, pembentukan T-DNA:AcvB kompleks secara in vitro dipelajari dengan menggunakan teknik imunopresipitasi (gb. 25). • ssT-DNA (2 kb) mula-mula diinkubasi dalam campuran reaksi yang mengandung protein AcvB. Campuran reaksi yang telah diinkubasi tersebut kemudian diimunopresipitasi baik dengan serum anti-AcvB maupun dengan serum preimun dan dianalisa dengan Southern blot analysis menggunakan probe T-DNA (probe 4). • T-DNA terdeteksi pada saat ssT-DNA diinkubasi dengan protein AcvB dan diimunopresipitasi dengan serum anti-AcvB, namun tidak ditemukan ketika diinkubasi dengan menggunakan serum preimun atau ketika protein AcvB tidak terdapat dalam reaksi (gb. 25). • Hasil ini mengindikasikan bahwa T-DNA:AcvB kompleks terbentuk secara in vitro.
4.3.4 Pendeteksian T-strand Kompleks yang Mengandung Protein AcvB dan VirE2 dalam Periplasma A. tumefaciens yang Terinduksi Acetosyringone (As). • Seperti yang telah disebutkan sebelumnya, AcvB menunjukkan adanya aktivitas ikatan ssDNA secara in vitro. • Dan AcvB dilokalisasi dalam periplasma A.tumefaciens (45). Dengan demikian, telah diuji apakah T-strand:AcvB kompleks terdapat dalam periplasma sel A. tumefaciens yang terinduksi As atau tidak (gb. 26). • Fraksi protein periplasmik disiapkan baik dari sel A. tumefaciens yang terinduksi As maupun yang tidak terinduksi As. • Fraksi-fraksi periplasmik tersebut diimunopresipitasi dengan serum anti-AcvB dan selanjutnya diuji dengan analisis Western dan Southernblot. • Dalam analisis Western blot, protein yang terimobilisasi pada membran diwarnai secara imunologi (immuno-stained) dengan menggunakan sera anti-AcvB dan anti-VirE2. • Dalam analisis Southern blot, DNA dideteksi dengan menggunakan probe T-DNA (probe 4).
Protein AcvB terdeteksi dalam strain A208 baik yang terinduksi As maupun yang tak terinduksi (panel A, lane 4 dan 5). • Hasil ini sesuai dengan penelitian sebelumnya yang mengungkapkan bahwa gen acvB diekspresi secara teratur dalam A. tumefaciens (45). • Dalam strain A208 yang terinduksi As, baik protein VirE2 maupun T-strand teramati (panel B, lane 3 dan panel C, lane 1). • Di sisi lain, baik protein VirE2 maupun T-strand tidak terdeteksi dalam strain A208 yang tidak terinduksi As (panel B, lane 1; panel C, lane 2). • Hasil tersebut mengindikasikan bahwa T-strand kompleks yang mengandung protein AcvB dan VirE2 terdapat dalam periplasma strain A208 yang terinduksi As (acvB+). Sedangkan, kompleks tersebut tidak terdapat pada periplasma strain A208 yang tidak terinduksi As atau dalam strain B119 (acvB -) baik yang terinduksi As ataupun yang tidak terinduksi.
4.3.5 Stimulasi Penggabungan ssDNA ke Dalam Protoplas Daun Tomat oleh Protein AcvB • [32P ssDNA] (probe 3) dipre-inkubasi dengan protein AcvB dan kemudian digabungkan ke dalam protoplas daun tomat. • Time courseanalysis mengindikasikan bahwa ssDNA:AcvB kompleks tergabung secara linier ke dalam protoplas selama 60 menit dan kemudian mencapai kondisi stabil (plateu), sementara ssDNA sendiri tidak tergabung secara signifikan (gb. 27). • Berdasarkan hasil ini, penggabungan ssDNA:protein kompleks ke dalam protoplas dibandingkan setelah 60 menit inkubasi dalam percobaan selanjutnya. • Aktivitas stimulasi penggabungan protein AcvB dibandingkan dengan protein lainnya (gb. 28). • Baik protein AcvB maupun B.S.A. yang direbus tidak menstimulasi penggabungan ssDNA ke dalam protoplas daun tomat.
Percobaan tersebut dilakukan untuk menentukan apakah ssDNA:AcvB kompleks masih berada dalam bentuk kompleks dalam protoplas. • Dalam percobaan ini, diasumsikan bahwa ssDNA:AcvB d kompleks dapat diimunopresipitasi dengan serum anti-AcvB, sedangkan ssDNA bebas tidak. • [32P] T-DNA (probe 4) dipre-inkubasi dengan protein AcvB selama 30 menit dan kemudian digabungkan dalam protoplas selama 1 jam. • Selanjutnya, protoplas tersebut dikacaukan dan fraksi supernatannya (sitosol) dikumpulkan dengan sentrifugasi pada 19 x g selama 10 menit. • Fraksi tersebut diimunopresipitasi dengan serum anti-AcvB dan radioaktivitas dalam immunopresipitatnya dihitung. • Hasilnya menunjukkan bahwa 83% [32P] ssDNA dalam sitosol terdapat dalam bentuk ssDNA:AcvB kompleks.
4.3.6 Hambatan Penggabungan ssDNA ke dalam Protoplas Daun Tomat dengan Serum Anti-AcvB • [32P] ssDNA (probe 3) dipre-inkubasi dengan AcvB baik bersama serum anti-AcvB maupun serum preimun dan kemudian digabungkan ke dalam protoplas daun tomat (gb. 29). • Penggabungan ssDNA yang telah dipre-inkubasi dengan adanya serum anti-AcvB terhambat secara signifikan dibandingkan dengan ssDNA yang dipre-inkubasi dengan adanya serum preimun atau tanpa serum. • Karenanya, hal ini menunjang hipotesis kami bahwa pembentukan T-DNA:AcvB kompleks membantu proses penggabungannya ke dalam protoplas.
4.3.7 T-DNA Tidak Hanya Terserap pada Permukaan Protoplas Melainkan Tergabung dalam Protoplas • Tiga percobaan dilakukan untuk menunjukkan bahwa ssDNA tidak hanya sekedar terserap pada permukaan protoplas tapi betul-betul tergabung dalam protoplas (percobaan pada gb. 27 dan 28). • Dalam percobaan ini, digunakan probe 4, bukan probe 3, karena probe 4 memiliki radioaktivitas spesifik yang lebih tinggi daripada probe 3, dan oleh karena itu, diharapkan bisa memberikan sensitivitas yang lebih tinggi dalam pengujiannya. • Pada percobaan pertama (gb. 30), protoplas yang telah ditambahkan kompleks probe 4:AcvB sebelumnya, dicuci secara merata dan kemudian diinkubasi dalam rentang waktu berbeda bersama proteinase K maupun DNase.
Dalam waktu inkubasi manapun, baik protoplas yang diinkubasi dengan adanya proteinase K maupun DNase dapat menahan radioaktivitas dibandingkan dengan protoplas yang diinkubasi tanpa enzim. • Selain itu, seluruh protoplas dapat menahan sebanyak 85% dari radioaktivitas awal setelah inkubasi selama 60 menit. • Pada percobaan kedua, protoplas yang ditambahkan probe4:AcvB kompleks sebelumnya dicuci secara merata, dan kemudian dipecah dengan menggunakan homogeniser teflon. • Homogenate disentrifugasi pada 19 k x g selama 10 menit untuk memisahkan supernatan yang mengandung sitosol dan endapannya yang mengandung membran plasma. • Jika [32P] ssDNA (probe 4) diserap pada permukaan protoplas, radioaktivitasnya sebagian besar akan terdapat dalam pelet. Sebaliknya, sekitar 67% dari probe 4 terdeteksi dalam supernatan (gb. 31A).
Hasil yang serupa didapatkan saat probe 3 digunakan dalam percobaan (gb. 31B). • Pada percobaan ketiga, protoplas yang ditambahkan T-DNA (2kb):AcvB kompleks sebelumnya, diteliti ada tidaknya protein AcvB di dalamnya dengan menggunakan teknik imunofluorescence tak langsung. • Protoplas yang diwarnai secara immunologi (immuno-stained) dipotong secara optik dengan menggunakan laser confokal microskope. • Gambar potongan tersebut menunjukkan bahwa protein AcvB tergabung ke bagian dalam protoplas (gb. 32). • Seluruh hasil yang yang disebutkan di atas menunjukkan secara pasti bahwa ssDNA:AcvB kompleks tergabung ke dalam protoplas.
4.3.8 Arah Tujuan T-DNA:AcvB Kompleks dalam Protoplas Daun Tomat • Sudah diketahui secara luas bahwa T-strand (atau T-kompleks) ditransfer ke dalam inti sel tanaman inang dan akhirnya tergabung ke dalam genom. Sehingga, perlu diteliti apakah AcvB sendiri mampu mengarahkan ssT-DNA ke dalam inti. • Protoplas diisi dengan T-DNA [32P]:AcvB kompleks dan kemudian diinkubasi selama 5 jam. • Dalam rentang waktu inkubasi yang beragam, protoplas dikacaukan dan disentrifugasi untuk memisahkan fraksi endapan inti (nuclear) dan supernatannya yang mengandung sitosol dan organela lainnya yang memiliki kerapatan sel lebih rendah. • Fraksi inti (nuclear) yang disiapkan dalam studi ini diharuskan tidak begitu tercemar oleh komponen lain yang berasal dari distribusi beberapa enzim penanda, klorofil dan DNA (Tabel 6). • Distribusi [32P] T-DNA dalam fraksi endapan inti (nuclear) dan supernatan tidak berubah secara signifikan melalui periode inkubasi (gb. 33). Hasilnya mengindikasikan bahwa T-DNA:AcvB kompleks tidak ditranslokasi ke dalam inti (nucleus).