Download

1 / 19
200 likes | 380 Views
Chap. 7 Transcriptional Control of Gene Expression (Part B). Topics RNA Polymerase II Promoters and General Transcription Factors Regulatory Sequences in Protein-coding Genes and the Proteins Through Which They Function. Goals

E N D
Chap. 7 Transcriptional Control of Gene Expression (Part B) • Topics • RNA Polymerase II Promoters and General Transcription Factors • Regulatory Sequences in Protein-coding Genes and the Proteins Through Which They Function • Goals • Learn about transcription control elements and methods of promoter analysis. • Learn how the RNA polymerase II pre-initiation complex assembles at a promoter. • Learn about the structures of eukaryotic transcription factors. Transcriptionally active polytene chromosomes
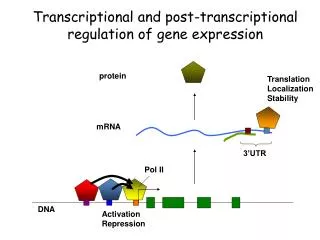
Overview of Eukaryotic Promoters The promoter of a eukaryotic gene can be defined as a sequence that sets the transcription start site for RNA polymerase. Strong RNA Pol II promoters contain an A/T rich sequence known as the TATA box located 26-31 bp upstream of the start site (Fig. 7.14). Other genes have alternative sequence elements known as initiators (Inr) which also serve as promoters that set the RNA Pol II start site. Finally, CG-rich repeat sequences (CpG islands) are used by RNA Pol II as promoters in 60-70% of genes. Most of these genes are weakly expressed.
Transcription Initiation by RNA Pol II RNA Pol II requires general TFs in addition to tissue-specific transcription factors for transcription of most genes in vivo. General TFs position RNA Pol II at start sites and assist the enzyme in melting promoter DNA. General TFs are highly conserved across species. The general TFs used at TATA box promoters are TFIIA, B, D, etc. TFIIA is required for transcription only in vivo. TFIID consists of TBP (TATA box binding protein) and 13 TBP-associated factors (TAFs). While the complete TFIID complex is required for transcription in vivo, only TBP is required in vitro. Formation of the pre-initiation complex in vitro is illustrated in the next two slides (Fig. 7.17).
Pol II Pre-initiation Complex Formation (I) The sequential steps leading to the assembly of the RNA Pol II pre-initiation complex in vitro are shown in Fig. 7.17. First, TBP binds to the TATA box and bends DNA near the promoter. Next, TFIIB binds, and then a complex between Pol II and TFIIF loads onto the promoter. TFIIF positions the Pol II active site at the mRNA start site. TFIIF also possesses histone acetylase activity and helps maintain chromatin at the promoter in an uncondensed state. TFIIE then binds creating a TFIIH docking site (next slide).
Pol II Pre-initiation Complex Formation (II) With the addition of TFIIH, the assembly of the pre-initiation complex is complete. Subsequently, one subunit of TFIIH melts DNA at the promoter, obtaining energy by ATP hydrolysis. Pol II then begins transcribing the mRNA. Another subunit of TFIIH phosphorylates the Pol II CTD, making Pol II highly processive. In vitro, all factors except TBP dissociate from the promoter region after Pol II moves downstream. Tissue-specific TFs bound to enhancers and promoter-proximal elements also play important roles in transcription initiation in vivo.
Linker Scanning Mutagenesis Analysis of Gene Regulatory Sites A technique called linker scanning mutagenesis commonly is used to identify transcription control regions known as promoter-proximal elements that lie within 100-200 bp of a start site Fig. 7.21. These elements are required for transcription but are not directly involved in start site selection. Large changes in the locations of these elements can interfere with transcription. Promoter-proximal elements are commonly important for cell type-specific transcription of genes.
Summary of Gene Control Elements A spectrum of control elements regulate transcription by RNA Pol II in eukaryotes. Their locations relative to the exons of a gene are summarized in Fig. 7.22. Enhancers are transcription control elements of 50-200 bp in length that can act from sites distant from the regulated gene. They often are important for cell type-specific regulation of transcription. Enhancers can be positioned upstream, downstream, or even within introns while still being functional. They further may be located 50 kb away from a transcription start site. Enhancers are composed of ~6-10 bp DNA modules that are bound by transcription factors. Promoter-proximal elements typically need to be with ~200 bp of the transcription start site to be functional. Yeast genes usually contain only upstream activating sequences (UAS) and a TATA box for control of transcription. UASs act similarly to enhancers and promoter-proximal elements in higher eukaryotes.
DNase I Footprinting The human genome encodes ~2,000 transcription factors (TFs). A method (DNase I footprinting) for determining locations of TF binding sites in DNA is shown in Fig. 7.23. First, DNA labeled on one strand is incubated with the protein of interest. Then the complex is treated with a small amount of DNase I, which cleaves DNA where it is not masked by the TF (Fig. 7.23a). A control DNA sample lacking the TF is treated under parallel conditions. Finally, the banding patterns from the two samples are compared by gel electrophoresis to locate the "footprint" region where the TF has shielded the DNA from cleavage (Fig. 7.23b).
Analysis of TF Activity in vivo TFs can be assayed for their ability to bind to DNA control elements and regulate gene expression by in vivo transfection assays (Fig. 7.25). In this method, a plasmid encoding the putative TF (protein X) is introduced into an animal cell along with a second vector encoding a reporter gene and the putative protein X binding site. If protein X binds to the site and is a transcription activator, then the reporter gene is switched on. Note that the cells must not express protein X per se.
Modular Structure of Activators I Transcription activators are modular proteins composed of distinct functional domains. They typically contain both DNA-binding and activation domains. A deletion analysis performed with the yeast GAL4 activator illustrating that it contains these two types of domains is shown in Fig. 7.26. The N-terminal amino acids of GAL4 modulate DNA binding, whereas its C-terminal region contains an activation domain.
Modular Structure of Activators II Functional domains in activators are joined by flexible protein linker sequences (Fig. 7.27). Due to the presence of linkers, the spacing and location of DNA control elements often can be shifted without interfering with DNA binding and regulation of promoters. The evolution of gene control regions through shuffling of DNA binding sequences between genes may have been favored due to the lack of strong requirements for control element spacing and location. The evolution of new activator protein genes through domain swapping has probably also been facilitated by linker sequences. Note, that transcription of some genes is controlled by repressors. Repressors typically contain a DNA-binding domain and a repression domain. The repression domain interacts with other TFs at a control site, inhibiting their activity. The inactivation of a repressor can lead to constitutive expression of the gene it controls.
Secondary Structure Motifs Secondary structure motifs are evolutionarily conserved collections of secondary structure elements which have a defined conformation. They also have a consensus sequence because the aa sequence ultimately determines structure. A given motif can occur in a number of proteins where it carries out the same or similar functions. Some well known examples such as the coiled-coil, EF hand/helix-loop-helix, and zinc-finger motifs are illustrated in Fig. 3.9. These motifs typically mediate protein-protein association, calcium/DNA binding, and DNA or RNA binding, respectively.
Helix-turn-helix TFs DNA-binding proteins bind specifically to DNA via non-covalent interactions. a-helices are one of the most common types of DNA-binding sequences (Fig. 7.28). The side-chains of residues within the a-helix often bind to the surfaces of bases exposed in the major groove of double-helical DNA. Binding to phosphates and bases in the minor groove typically is less important. One of the most common DNA-binding structure motifs is the helix-turn-helix.The second helix in this motif (the DNA recognition helix) typically binds to a specific sequence of bases in DNA. The recognition helices in the dimeric bacteriophage 434 repressor are indicated with asterisks in Fig. 7.28a. Helix-turn-helix TFs are common in bacteria.
Zinc-finger TFs The most common DNA-binding motif in human and multicellular animal TFs is the zinc finger. Two types of zinc finger TFs are discussed here--C2H2zinc finger TFs (Fig. 7.29a) and C4zinc finger TFs (Fig. 7.29b). Most TFs that contain C2H2 zinc fingers are monomeric. Its 2 cysteine and 2 histidine residues bind to zinc ions (Zn2+) (Fig. 7.29a), and the a-helix containing the 2 histidines binds to bases in the major groove. Much less common are TFs containing C4 zinc fingers. Most TFs containing this motif are dimeric. Nuclear receptors, which bind steroid hormones and other compounds, contain this motif. The glucocorticoid receptor is shown in Fig. 7.29b. Zinc ions are bound to the DNA recognition helix of this motif, which contacts bases in the major groove.
Leucine-zipper TFs Leucine-zipper TFs contain extended a-helices wherein every 7th amino acid is leucine. This periodicity creates a nonpolar face on one side of the helix that is ideal for dimerization with another such protein via a coiled-coil motif (Fig. 7.29c). So-called basic zipper (bZip) TFs have a similar structure except that some leucines are replaced by other nonpolar amino acids. The N-terminal ends of both leucine-zipper and bZip proteins contain basic amino acids that interact with bases in the major groove (Fig. 7.29c). Leucine zipper proteins are now considered to be a subclass of bZip proteins. Another class of TF, the basic helix-loop-helix (bHLH) proteins are similar to bZip proteins, but contain a loop between the DNA recognition helix and the coiled-coil region (Fig. 7.29d). bZip and bHLH proteins commonly form heterodimeric TFs. Basic residues
Regulation of TF Activity Many TFs bind ligands or co-activator/co-repressor proteins that modulate their structure and activity. In the yeast GAL4 TF, its "acidic activation domain" adopts an essentially random-coil structure until it binds to a co-activator protein. This control mechanism keeps the TF turned off until the appropriate cofactor is present in the nucleus. Nuclear receptors such as the estrogen receptor contain partially structured activation domains that undergo conformational changes to the active structural state on binding to hormone (e.g., estrogen) (Fig. 7.30b). In the active conformation, the estrogen receptor can bind to co-activator proteins required for transcription. The estrogen antagonist, tamoxifen, that is used in breast cancer therapy, locks the receptor in its inactive conformation that cannot bind co-activator proteins (Fig. 7.30c).
Heterodimeric TFs The formation of heterodimeric TFs by bZip and bHLH TFs is important in increasing the complexity of transcriptional regulation of genes. Sometimes monomers within the heterodimer recognize the same DNA element, but have different activation domains (Fig. 7.31a). Different regulatory responses result from the different combinations of activation domains bound to the site. In other cases, monomers within the heterodimer bind different DNA elements (Fig. 7.31b). Each site then binds a unique species of heterodimer. Lastly, an inhibitory factor that binds to only one type of monomer will only affect sites used by that monomer (Fig. 7.31c).
Cooperative Binding of TFs to DNA In many cases, a TF will bind to a DNA element with high affinity only when complexed with a second TF (Fig. 7.32a). Such cooperative binding to DNA adds further complexity to gene regulation. Namely, a certain TF will bind to its DNA element only if its interacting partner also is expressed in that cell type. In addition, expression levels of interacting TFs can be varied between tissues to adjust gene transcription rates.
Multiprotein Complexes at Enhancers Enhancers typically contain several DNA sequence elements that are recognized by different DNA binding proteins (e.g., the ß-interferon enhancer, Fig. 7.33). The resulting nucleoprotein complex is called an enhanceosome. Enhancers often are involved in tissue-specific control of transcription. Given the complex structures of enhanceosomes, it is easy to see how the absence of even one of the factors that bind to the enhancer in a certain tissue could change the expression level of the gene.





![[VI]. Post-Transcriptional Processing and Post-Transcriptional Control of Gene Expression](https://cdn1.slideserve.com/3214110/vi-post-transcriptional-processing-and-post-transcriptional-control-of-gene-expression-dt.jpg)
![[V]. Process of Transcription and Transcriptional Control of Gene Expression](https://cdn2.slideserve.com/5058527/v-process-of-transcription-and-transcriptional-control-of-gene-expression-dt.jpg)