Download
1 / 43
430 likes | 513 Views
Explore the structure, properties, and mechanisms of various transposon elements in DNA and their roles in chromosomal rearrangement and genetic evolution.

E N D
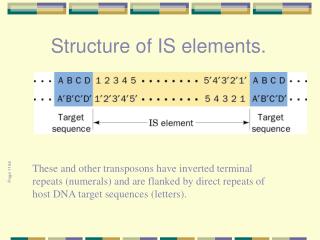
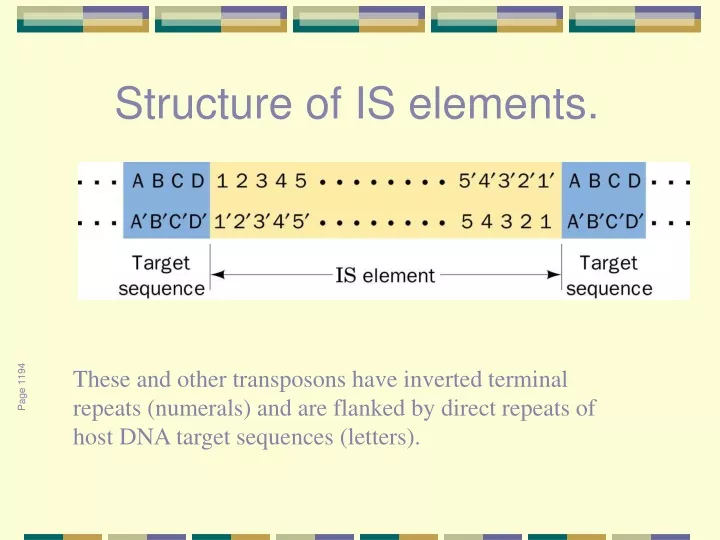
Structure of IS elements. These and other transposons have inverted terminal repeats (numerals) and are flanked by direct repeats of host DNA target sequences (letters). Page 1194
A model for the generation of direct repeats of the target sequence by transposon insertion. Page 1195
Properties of Some Insertion Elements. Page 1194
A map of transposon Tn3. Page 1195 Total size 4957 bp. Inverted terminal repeats 38 bp each
A composite transposon. Page 1195 The IS-like modules may have either (a) direct or (b) inverted relative orientations.
Electron micrograph of a single-stranded circular DNA containing a transposon. Page 1215
The cut-and-paste transposition mechanism catalyzed by Tn5 transposase. Page 1195
Comparison of transposition pathways. Grey segments represent transposable DNA elements. Small arrows indicate phosphodiester bond breakage. Solid and dotted lines represent donor and target DNA, respectively
X-Ray structure of Tn5 transposase. Page 1196 in complex with a 20-bp DNA containing the OE sequence
Replicative transposition. Page 1197 This type of transposition inserts a copy of the transposon at the target site while another copy remains at the donor site.
A cointegrate. Page 1197 This structure forms by the fusion of two plasmids, one carrying a transposon, such that both junctions of the original plasmid are spanned by transposons with the same orientation (arrows).
A model for transposition involving the intermediacy of a cointegrate. Here more lightly shaded bars represent newly synthesized DNA. Page 1197
Chromosomal rearrangement via recombination. Page 1199 (a) The inversion of a DNA segment between two identical transposons with inverted orientations.
Chromosomal rearrangement via recombination.. (b) The deletion of a DNA segment between two identical transposons with the same orientation
The mechanism of phase variation in Salmonella. hin codes for Hin DNA invertase hix : Two sites, closely related, 26 bp (2 x 12 bp imperfect inverted repeats separated by 2 bp) H1, H2: Genes for two antigenically distinct flagellin proteins rh1: Gene for H1 gene repressor Page 1200
Repetitive DNA interspersed in tandem I
Moderately Repetitive Sequences in the Human Genomea Page 1437
Elementer i det humane genom som kan transposeres på en RNA-formidlet måte
Alu elements • Length = ~300 bp • Repetitive: > 1,000,000 times in the human genome • Constitute >10% of the human genome • Found mostly in intergenic regions and introns • Propagate in the genome through retroposition (RNA intermediates).
Alu elements can be divided into subfamilies The subfamilies are distinguished by ~16 diagnostic positions.
Alu sequences in the globin gene cluster Page 1444
Gene sequences of (a) retroviruses and(b) the Ty1 retrotransposon from yeast. Page 1203
The catalytic mechanism of 5-methylcytosine methyltransferases (m5C-MTases). Page 1205
X-Ray structure of M.HhaI Page 1206 in complex with S-adenosylhomocysteine and a duplex 13-mer DNA containing a methylated f5C residue at the enzyme’s target site.
Maintenance methylation. Page 1207
CpG-frekvens og CpG-øyer The typical density of CpG doublets in mammalian DNA is ~1/100 bp, as seen for a -globin gene. In a CpG-rich island, the density is increased to >10 doublets/100 bp. The island in the APRT gene starts ~100 bp upstream of the promoter and extends ~400 bp into the gene. Each vertical line represents a CpG doublet.
Vedlikeholdsmetylering Ved maintenance-metylering induserer metyleringsmønsteret i en parental DNA-tråd det tilsvarende metyleringsmønster i den komplementære tråden. Slik kan et stabilt metyleringsmønster opprettholdes i en cellelinje
CpG – underrepresentert i genomet The CpG doublet occurs in vertebrate DNA at only ~20% of the frequency that would be expected from the proportion of G·C base pairs. (this is because CpG doublets are methylated on C, and spontaneous deamination of methyl-C converts it to T, introducing a mutation that removes the doublet.) In certain regions, however, the density of CpG doublets reaches the predicted value; in fact, it is increased by 10× relative to the rest of the genome. The CpG doublets in these regions are unmethylated
Evolusjon av CpG-øyer: en mulig mekanisme Ancestralt eukaryot genom med metylering av C i CpG, bortsett fra i visse genassosierte områder Metylerte CpG muteres gradvis til TpG eller CpA, mens umetylerte CpG forblir
TABLE 1 DISEASES OF TRINUCLEOTIDE REPEATS NAME OF THE DISEASE SEQUENCE OF THE REPEAT LOCATION OF THE REPEAT Fragile site 11B Fragile X syndrome CGG EXON Dentatorubral-pallidoluysian atrophy Haw river syndrome Huntington's disease Machado-Joseph disease Spinal and Bulbar muscular dystrophy Spinocerebellar ataxia type 1 CAG EXON Myotonic dystrophy CTG EXON Friedrich's ataxia GAA INTRON Trinucleotide expansion diseases
The loop-out mechanism for the alteration of the number of consecutive triplet repeats in DNA through its replication. Page 1209