Download
1 / 60
600 likes | 610 Views
Origins of Life – Another View (Lane) Foundational Elements: - 1. Eukaryotic life evolved when an archaeon engulfed a eubacterial proto-mitochondrion.

E N D
Origins of Life – Another View (Lane) • Foundational Elements: • - 1. Eukaryotic life evolved when an archaeon engulfed a eubacterial proto-mitochondrion. • - 2. In addition to shared DNA and protein synthesis machinery, the generation of energy by a ‘proton pump’ is common to all life AND non-random – a plesiomorphic adaptation. • - 3. The energetics of life, which achieve ordered states through increasing the entropy of the system, requires a continual source of energy.
II. Energetics First Law of thermodynamics:
II. Energetics First Law of thermodynamics: total energy of a system is constant; energy can neither be created nor destroyed
II. Energetics First Law of thermodynamics: total energy of a system is constant; energy can neither be created nor destroyed Second law:
II. Energetics First Law of thermodynamics: total energy of a system is constant; energy can neither be created nor destroyed Second law: entropy of an isolated system always increases over time “X” entropy “<X” entropy
II. Energetics Chemical Reactions: There are two stable states here; of the reactants and the product. The product is at a lower energy state, so the reaction will occur spontaneously if energy is ‘added’ to the reactants. Energy will be released as heat, increasing the total entropy of the system – even though the products may appear to be matter in a “more ordered” form.
II. Energetics Chemical Reactions: The “ordered” form of lipids and proteins in a spherical double-membrane is a good example of molecules achieving a stable, though “ordered” state.
II. Energetics Chemical Reactions: The “ordered” form of a polypeptide, also, has a lower energy state than free amino acids – but they must be activated to bond.
II. Energetics Chemical Reactions: Activation will occur spontaneously if there are reactive precursors… The “ordered” form of a polypeptide, also, has a lower energy state than free amino acids – but they must be activated to bond.
Reactive precursors will form if there is a reactive environment. Activation will occur spontaneously if there are reactive precursors… The “ordered” form of a polypeptide, also, has a lower energy state than free amino acids – but they must be activated to bond.
Life’s complexity arises as a consequence of energy harvest from the environment, PUSHING energetic reactions that give off heat and increase entropy. Life HEAT ENERGY LOST
Life’s complexity arises as a consequence of energy harvest from the environment, PUSHING energetic reactions that give off heat and increase entropy. A continuous energy flux is required to maintain and grow this system in an oxidizing atmosphere that would drive the reactions the other way. Life HEAT ENERGY LOST
Life’s complexity arises as a consequence of energy harvest from the environment, PUSHING energetic reactions that give off heat and increase entropy. A continuous energy flux is required to maintain and grow this system in an oxidizing atmosphere that would drive the reactions the other way. Also, the concentration of reactants must be greater than the concentration of products to drive the reaction in this direction. Life HEAT ENERGY LOST
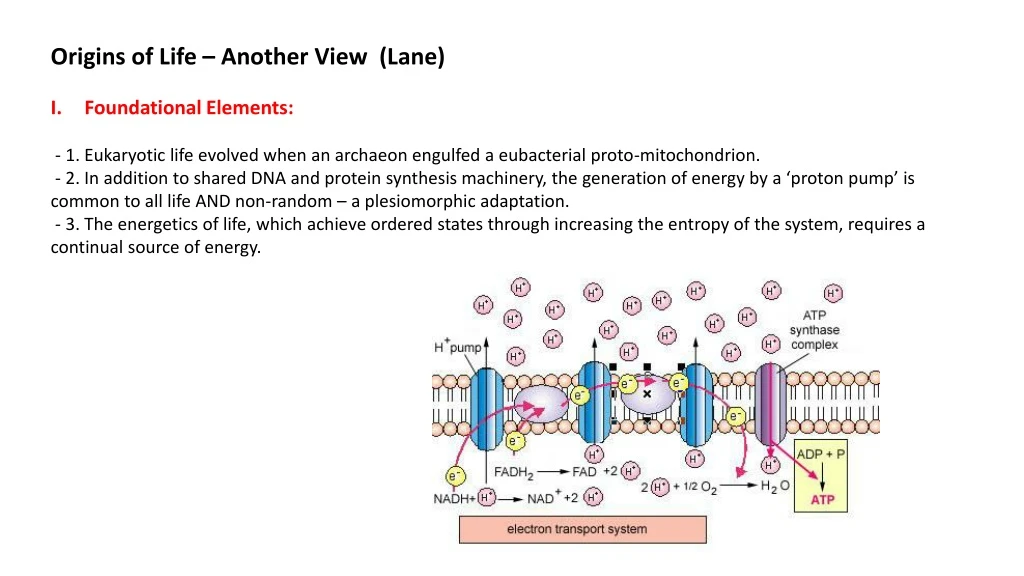
II. Energetics Chemical Reactions: Redox reactions – passing electrons between atoms/molecules. Electrons are stripped from food and passed down an electron transport chain of charged iron (Fe+3) in iron-Sulphur clusters to establish a proton gradient. The flow of protons is used to make ATP, and the original electron is ultimately passed to O2, making water. For each pair of electrons passed, 10 H+ ions cross the membrane 1021 protons pumped in your body every second.
II. Energetics Chemical Reactions: Charge differential of 150-200millivolts, across a very thin membrane (6 nm)… field strength = 30 million volts/m … lightening bolt… 1000 x capacity of household wiring. Maintaining this differential requires a membrane impermeable to H+ ions, so their flow can be regulated by proteins.
II. Energetics Chemical Reactions: ATP Synthase: For every 10 H+, the rotor spins once and releases 3 ATP. Head can spin 100x per second.
II. Energetics Electron Transport Chains (Proton Pumps): e- e- “FOOD” O2
II. Energetics Electron Transport Chains (Proton Pumps): e- e- “FOOD” O2 Any electron Donor: NADH, FADH, H2, H2S, Fe+2, H2O Any Oxidant: O2, NO2, NO3, SO2, SO3, CO2 These redox pathways include photosynthetic pathways, obviously.
After 4 billion years of evolution, life has evolved enzymes that are efficient by reducing the activation energy and facilitating these redox reactions. Even with this efficiency, however, there is often a greater loss of energy (40x) as entropy than energy stored in life. Life 40x = HEAT ENERGY LOST
The earliest life forms/reactions must have been more inefficient – so to produce the same products, as a smaller fraction of energy, there must have been lots more energy pumping through the system. Life
III. An Alternative Model A. Problems with the RNA World Model Watson and Crick Miller and Urey Monomers Functional RNA “warm little pond”
III. An Alternative Model A. Problems with the RNA World Model Watson and Crick Miller and Urey Monomers Requires source of continuous environmental energy AND carbon, and continual production of activated nucleotides to drive the reactions to form RNA. Functional RNA “warm little pond” Lightening and other sources of energy do not provide the carbon, and they are unstable and destructively high energy
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative What is Needed to Build a Cell? Continuous flow of energy Continuous flow of carbon as source of new molecules Rudimentary catalysts (could be inorganic FeS minerals that are often at the heart of organic catalysts) “Focusing” products to raise their concentrations to reactive levels Outflow of products and waste “Life as a Dissipative Structure” – control and regulating the flow of energy and matter to produce something Life
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative Hydrothermal Vents “Black Smokers” Very hot – 250-400oC, driven by magma in contact with water. Most stable carbon compound is CO2. Very unstable, collapsing in decades Do have lots of FeS compounds but precipitated organics would be blown from chimney.
Hydrothermal Vents “White Smokers” of Today Mantle rock near spreading centers is exposed to sea water; the olivine (rich in ferrous iron) is oxidized by water in an exothermic reaction, giving off heat and H2. The warm (60-90oC), H-rich water percolates up and the carbonate minerals precipitate into microporous chimneys. High flux of carbon and energy is channeled over inorganic catalysts Thermal currents through pores can concentrate organics 1000 – 106 fold Vents persist for millennia 500 um
Hydrothermal Vents “White Smokers” of the Hadean (4bya) Oceans more acidic (more CO2 in atmosphere, more dissolved in water). CO2 remains in suspension with H2, not precipitating out as carbonate. Either WON’T react, or will proceed to to CH4… but that’s not good for life. Need it to react to intermediates like formaldehyde (CH2O). At pH = 7, If H2 gives up it’s electrons to CO2, the formate will pass the electrons right back to H+, giving CO2 and H2! But methanogens live on this reaction… so how do they do it? H2 2H+ + 2 e- CO2-2
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative: Metabolism First Pore Wall Percolating from mantle: pH = 11 OH H2 Seawater percolating through pores: pH = 6 H+ FeS STEP 1: The role of a proton gradient: If the formate (CO2- ) can accept a H+ to balance the charge, it won’t get rid of the electron. If the CO2 is placed in an acidic environment rich in H+, this will tend to happen. Likewise, if the free H+ is placed in an alkali environment (rich in OH-), it will bind to form stable water. The H2 has reacted with the CO2 irreversibly. If these fluids are separated by a thin inorganic membrane with FeS minerals, the FeS will draw the electrons from the H2 and transfer them to CO2. Physical structure of vent drives the synthesis of organics. These accumulate 1000x H2O CO2-2+ 2H+ CH2OO FeS 2e- 2 H+
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 2: Energy Harvest The proton gradient can be exploited by protein synthesized organically for Energy Harvest (ATP Synthase). (This presumes the evolution of a genetic system to encode ATP synthase… the genetic system has evolved as a consequence of the accumulation and synthesis of organics) So, both E harvest AND organic production ATP
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 2: Energy Harvest The proton gradient can be exploited by protein synthesized organically for Energy Harvest (ATP Synthase). The key here is that the gradient is maintained because H+ ions coming in will bind with OH- entering from the alkaline vent fluid, forming water. This keeps [H+] low in “cell”. Also, transport through protein favored over transport through inorganic barrier or protomembrane. So, both E harvest AND organic production ATP
How to “leave the nest”? But they would need to remain “leaky” to H+ to generate both power and organics. In this situation, they must stay in the vent. There is no selective value to evolving a modern “H+ pump”, because ions flow right back across the membrane. And there is no possibility of evolving a less permeable modern membrane without reducing OH- and losing gradient. “Chicken-egg” problem, getting to impermeable membrane with pump. ATP
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 3: Antiporters Methanogens actually pump out Na+, not H+! They have an “antiporter” (SPAP) that acts as a “revolving door”. When Na+ ions enter the cell in response to their concentration gradient, a H+ ion exits to balance the charge. But antiporters, like revolving doors, can move both ways. So, in our vent protocell, for each H+ that enters cell in response to its gradient, a Na+ would leave – establishing a Na+ gradient, too.
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 3: Antiporters Methanogens actually pump out Na+, not H+! Then, they have an “antiporter” (SPAP) that acts as a “revolving door”. When Na+ ions enter the cell in response to their concentration gradient, a H+ ion exits to balance the charge. But antiporters, like revolving doors, can move both ways. So, in our vent protocell, for each H+ that enters cell in response to its gradient, a Na+ would leave – establishing a Na+ gradient, too. For the LOVE OF GOD, why is this better????
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 3: Antiporters Membranes are much less permeable to Na+. So, even a membrane that is leaky to H+ can be impermeable to Na+… which means that Na+ can only re-enter the cell through a protein channel… Like ATP Synthase. (Which, actually, is what can happen in real methanogens… ATP synthase can be powered by a H+ or Na+ ion.) So, the cell still has a natural H+ gradient powering ATP production, but also a FREE Na+ gradient that can power ATP production – increasing ATP production by 60%. (The Na+ gradient is free because it is powered by the cost-free movement of H+ ions in response to their gradient.)
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 3: Antiporters In fact, almost all cells have enzymes that are optimized to working at LOW [Na+]… curious since cells evolved in an ocean rich in Na+. Maybe such a Na+ pump was ancestral, too. However, at this point, the cell is still dependent on the natural H+ gradient. (And again, with a leaky membrane, there is no benefit to evolving a H+ pump.)
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 4: Evolving the H+ Pump But with the antiporter, now a H+ pump is adaptive. Even with a membrane leaky to H+, those that are pumped out are more likely to come back across the anti-porter… so the WORK done to pump out H+ does establishes a gradient - a gradient of Na+. With this gradient, the flux of Na+ through ATP synthase is increased, ‘paying’ for the gradient.
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 4: Evolving the H+ Pump Likewise, a pump that pumps out Na+ increases the Na+ gradient…. This may change the direction of the anti-porter so that Na+ moves IN (in response to gradient), and H+ moves OUT (proto-H+ pump).
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 5: Evolving an impermeable membrane With a pump established, selection will favor protocells that evolve a progressively impermeable membrane that restricts ALL H+ and Na+ flow to proteins like ATP synthase.
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 5: Evolving an impermeable membrane With a pump established, selection will favor protocells that evolve a progressively impermeable membrane that restricts ALL H+ and Na+ flow to proteins like ATP synthase. And, with a pump AND impermeable membrane, the cell is no longer dependent on the natural H+ gradient. It can leave the vent and pump H+ ions to establish a gradient for ATP and carbohydrate production.
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative STEP 6: Divergence of Archaea and Bacteria Same genetic system – probably evolved during this process, selecting for membrane proteins and enzymes to synthesize membrane lipids. BUT: The types of lipids used could vary: Archea use one stereoisomer of glycerol and bacteria use another.
III. An Alternative Model A. Problems with the RNA World Model B. An Alternative
The Deep Divide: Same in Bacteria and Archaea: Chemiosmotic - Drive ATP synthesis with proton pumps Both have ATP synthase Ribosomes DNA, RNA, and protein synthesis Acetyl-CoA pathway of carbon fixation Different: Different enzymes for DNA replication Different lipids in cell membrane Were bacterial ancestral and Archaea evolved adaptations to extreme environments? No, even those in normal environments are different, and even bacteria in extreme environments are like other bacteria. r-RNA tree… one gene
Williams, et al. (2013) An archaeal origin of eukaryotes supports only two primary domains of life. Nature.
The Chimerism of Eukaryotes • How could this level of genetic relatedness with 25 groups of bacteria and 7-8 groups of Archaea arise? • Serial endosymbiosis or serial lateral gene transfer TO eukaryotes? It is too abrupt, all dating to origin of Eukaryotes. • Endosymbiosis of bacterial cell by archaean, and then divergence and LGT from the independent bacteria and archaeans among their clades.
III. An Alternative Model • A. Problems with the RNA World Model • B. An Alternative • C. Implications and Explanations • Why are eukaryotes the only clade to evolve multicellularity and structural complexity? • Eukaryotic cells are, on average, 15,000 larger in volume than prokaryotic cells. • Eukaryotic cells respire at 1/3 the rate (per unit volume), but thus harvest 5000x as much energy • (because they have 1000’s of mitochondria in this volume, harvesting energy). • 80% of the cells energy is used making proteins. • The more genes, the higher the cost of protein synthesis.
III. An Alternative Model • A. Problems with the RNA World Model • B. An Alternative • C. Implications and Explanations • Why are eukaryotes the only clade to evolve multicellularity and structural complexity? • Eukaryotic cells are, on average, 15,000 larger in volume than prokaryotic cells. • Eukaryotic cells respire at 1/3 the rate (per unit volume), but thus harvest 5000x as much energy • (because they have 1000’s of mitochondria in this volume, harvesting energy). • 80% of the cells energy is used making proteins. • The more genes, the higher the cost of protein synthesis. • Bacteria have ~5000 genes, eukaryotes have ~20,000. • So eukaryotes can distribute 5000x as much energy across 4x as many genes. • Eukaryotes, even with their larger genome, can spend 1200x as much energy / gene (making proteins). • If we expand the bacterial genome by 4x, to the size of a eukaryotic genome, then the energy / gene drops to 5000x less than eukaryotes. • So, eukaryotes can support a genome that is both larger, and 1000’sx more productive than prokaryotes. WHY?
III. An Alternative Model • A. Problems with the RNA World Model • B. An Alternative • C. Implications and Explanations • Why are eukaryotes the only clade to evolve multicellularity and structural complexity? • How about if we increase bacterial cell volume? Increasing r by 25x increases volume 15,000 fold. But it only increases SA 625 fold. Even then, protein synthesis needs to increase 625 fold to make 625x ATP synthase, just to keep pace with previous, “bacterial” levels of productivity. • The genome would have to increase 625 fold, just to keep the same ‘energy/gene’ ratio, to produce this 625-fold increase in protein demand. And this is just for SA proteins! The volume has increased 15,000 fold, and there are proteins needed in that huge, new space, too. • Energetically, bacteria are limited to being small. • The giant bacteria are the exceptions that prove the rule:
III. An Alternative Model • A. Problems with the RNA World Model • B. An Alternative • C. Implications and Explanations • Why are eukaryotes the only clade to evolve multicellularity and structural complexity? • The giant bacteria are the exceptions that prove the rule: • Epulopiscium – hind-gut of Surgeonfish – 0.5 mm long • (1000x large than E. coli) • - highly convoluted membrane increases SA for reactions. • - ‘extreme polyploidy’ – 200,000 copies of genome. • - shape – reduce V per unit SA • - still, about same energy/gene as E. coli, 5000x less than eukaryote.