Download
1 / 1
10 likes | 170 Views
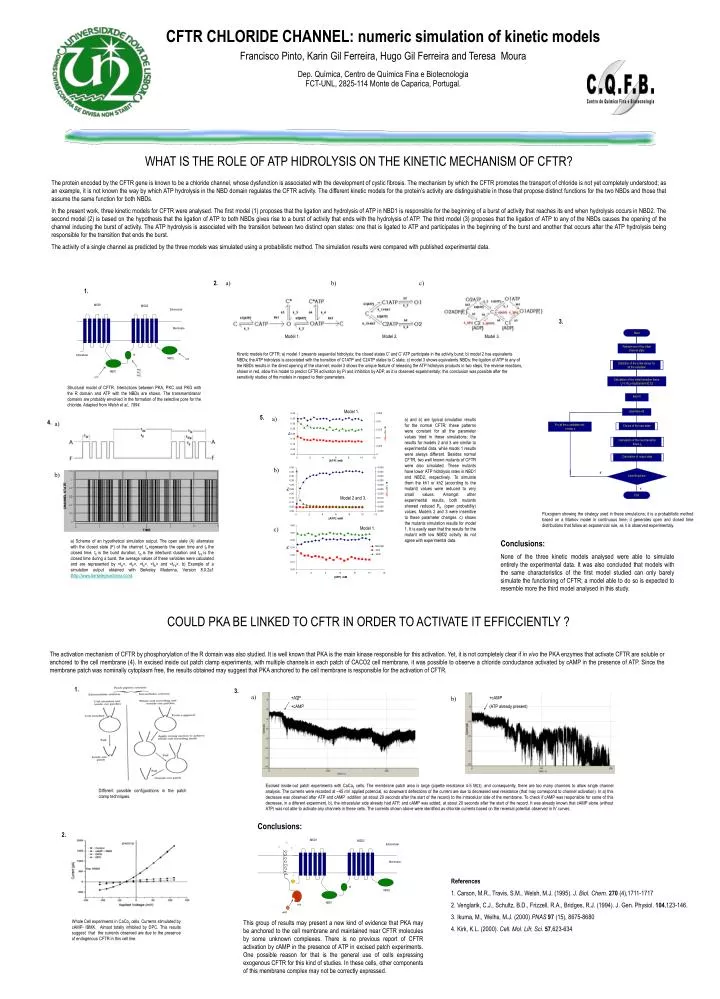
CFTR CHLORIDE CHANNEL: numeric simulation of kinetic models Francisco Pinto, Karin Gil Ferreira, Hugo Gil Ferreira and Teresa Moura Dep. Química, Centro de Química Fina e Biotecnologia FCT-UNL, 2825-114 Monte de Caparica, Portugal. Start. Random set of the initial channel state.

E N D
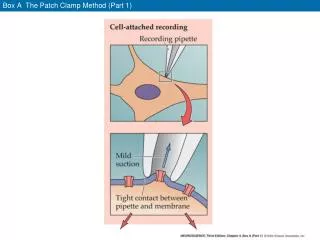
CFTR CHLORIDE CHANNEL: numeric simulation of kinetic models Francisco Pinto, Karin Gil Ferreira, Hugo Gil Ferreira and Teresa Moura Dep. Química, Centro de Química Fina e Biotecnologia FCT-UNL, 2825-114 Monte de Caparica, Portugal. Start Random set of the initial channel state Definition of the initial values for all the variables Calculation of the initial transition times: tij=-(1/kij)logn[random(0,1)] time=0 MSD1 MSD2 time=time+dt ? Extracellular ? ? MSD1 MSD2 Extracelular Membrane For all the x variables set: x=next x Membrane Choice of the next state R Intracellular NBD2 ? ATP PKA PKC PKG NBD1 ATP Model 1. Model 2. Model 3. R Calculation of the next transition times tij NBD2 Kinetic models for CFTR: a) model 1 presents sequential hidrolysis; the closed states C* and C* ATP participate in the activity burst; b) model 2 has equivalents NBDs; the ATP hidrolysis is associated with the transition of C1ATP and C2ATP states to C state; c) model 3 shows equivalents NBDs; the ligation of ATP to any of the NBDs results in the direct opening of the channel; model 3 shows the unique feature of releasing the ATP hidrolysis products in two steps, the reverse reactions, shown in red, allow this model to predict CFTR activation by Pi and inhibition by ADP, as it is observed experimentaly; this conclusion was possible after the sensitivity studies of the models in respect to their parameters. NBD1 Structural model of CFTR. Interactions between PKA, PKC and PKG with the R domain and ATP with the NBDs are shown. The transmembranar domains are probably envolved in the formation of the selective pore for the chloride. Adapted from Welsh et al., 1994. PKA cAMP Calculation of output data F time=final time V End Different possible configurations in the patch clamp techniques. WHAT IS THE ROLE OF ATP HIDROLYSIS ON THE KINETIC MECHANISM OF CFTR? The protein encoded by the CFTR gene is known to be a chloride channel, whose dysfunction is associated with the development of cystic fibrosis. The mechanism by which the CFTR promotes the transport of chloride is not yet completely understood; as an example, it is not known the way by which ATP hydrolysis in the NBD domain regulates the CFTR activity. The different kinetic models for the protein’s activity are distinguishable in those that propose distinct functions for the two NBDs and those that assume the same function for both NBDs. In the present work, three kinetic models for CFTR were analysed. The first model (1) proposes that the ligation and hydrolysis of ATP in NBD1 is responsible for the beginning of a burst of activity that reaches its end when hydrolysis occurs in NBD2. The second model (2) is based on the hypothesis that the ligation of ATP to both NBDs gives rise to a burst of activity that ends with the hydrolysis of ATP. The third model (3) proposes that the ligation of ATP to any of the NBDs causes the opening of the channel inducing the burst of activity. The ATP hydrolysis is associated with the transition between two distinct open states: one that is ligated to ATP and participates in the beginning of the burst and another that occurs after the ATP hydrolysis being responsible for the transition that ends the burst. The activity of a single channel as predicted by the three models was simulated using a probabilistic method. The simulation results were compared with published experimental data. 2. a) b) c) 1. 3. Model 1. 5. a) a) and b) are typical simulation results for the normal CFTR; these patterns were constant for all the parameter values tried in these simulations; the results for models 2 and 3 are similar to experimental data, while model 1 results were always different. Besides normal CFTR, two well known mutants of CFTR were also simulated. These mutants have lower ATP hidrolysis rates in NBD1 and NBD2, respectively. To simulate them the kh1 or kh2 (according to the mutant) values were reduced to very small values. Amongst other experimental results, both mutants showed reduced PA (open probability) values. Models 2 and 3 were insenitive to these parameter changes. c) shows the mutants simulation results for model 1. It is easily seen that the results for the mutant with low NBD2 activity do not agree with experimental data. 4. a) b) b) Model 2 and 3. Fluxogram showing the strategy used in these simulations; it is a probabilistic method based on a Markov model in continuous time; it generates open and closed time distributions that follow an exponencial rule, as it is observed experimentaly. c) Model 1. a) Scheme of an hypothetical simulation output. The open state (A) alternates with the closed state (F) of the channel. tA represents the open time and tF the closed time. tb is the burst duration, tib is the interburst duration and tFb is the closed time during a burst, the average values of these variables were calculated and are represented by <tA>, <tF>, <tb>, <tib> and <tFb>. b) Example of a simulation output obtained with Berkeley Madonna, Version 8.0.2a1 (http://www.berkeleymadonna.com). Conclusions: None of the three kinetic models analysed were able to simulate entirely the experimental data. It was also concluded that models with the same characteristics of the first model studied can only barely simulate the functioning of CFTR; a model able to do so is expected to resemble more the third model analysed in this study. COULD PKA BE LINKED TO CFTR IN ORDER TO ACTIVATE IT EFFICCIENTLY ? The activation mechanism of CFTR by phosphorylation of the R domain was also studied. It is well known that PKA is the main kinase responsible for this activation. Yet, it is not completely clear if in vivo the PKA enzymes that activate CFTR are soluble or anchored to the cell membrane (4). In excised inside out patch clamp experiments, with multiple channels in each patch of CACO2 cell membrane, it was possible to observe a chloride conductance activated by cAMP in the presence of ATP. Since the membrane patch was nominally cytoplasm free, the results obtained may suggest that PKA anchored to the cell membrane is responsible for the activation of CFTR. 1. 3. a) +ATP +cAMP b) +cAMP (ATP already present) + Excised inside-out patch experiments with CaCo2 cells. The membrane patch area is large (pipette resistance 4-5 M), and consequently, there are too many channels to allow single channel analysis. The currents were recorded at –45 mV applied potencial, so downward deflections of the current are due to decreased seal resistance (that may correspond to channel activation). In a) this decrease was observed after ATP and cAMP addition (at about 20 seconds after the start of the record) to the intracelular side of the membrane. To check if cAMP was responsible for some of this decrease, in a diferent experiment, b), the intracelular side already had ATP, and cAMP was added, at about 20 seconds after the start of the record. It was already known that cAMP alone (without ATP) was not able to activate any channels in these cells. The currents shown above were identified as chloride currents based on the reversal potential observed in IV curves. Conclusions: 2. References 1. Carson, M.R., Travis, S.M., Welsh, M.J. (1995). J. Biol. Chem.270 (4),1711-1717 2. Venglarik, C.J., Schultz, B.D., Frizzell, R.A., Bridges, R.J. (1994). J. Gen. Physiol. 104,123-146. 3. Ikuma, M., Welhs, M.J. (2000) PNAS97 (15), 8675-8680 4. Kirk, K.L. (2000). Cell. Mol. Lifr. Sci.57,623-634 Whole Cell experiments in CaCo2 cells.Currents stimulated by cAMP- IBMX. Almost totally inhibited by DPC. This results suggest that the currents observed are due to the presence of endogenous CFTR in this cell line. This group of results may present a new kind of evidence that PKA may be anchored to the cell membrane and maintained near CFTR molecules by some unknown complexes. There is no previous report of CFTR activation by cAMP in the presence of ATP in excised patch experiments. One possible reason for that is the general use of cells expressing exogenous CFTR for this kind of studies. In these cells, other components of this membrane complex may not be correctly expressed.