Download
1 / 28
290 likes | 453 Views
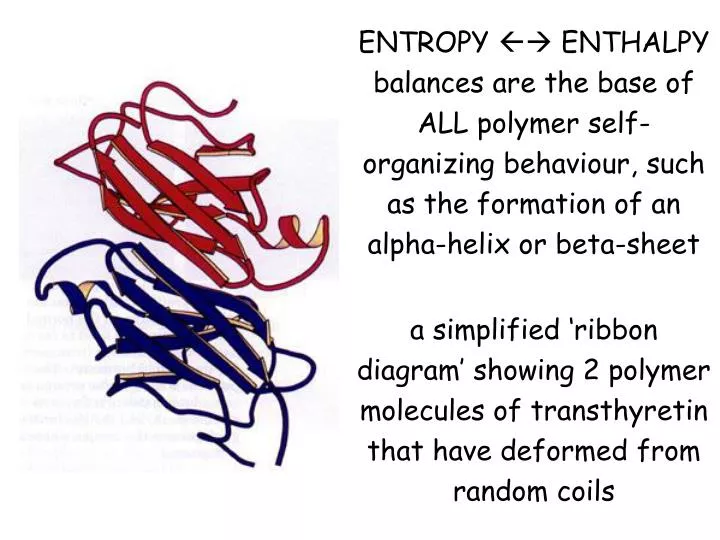
ENTROPY ENTHALPY balances are the base of ALL polymer self-organizing behaviour, such as the formation of an alpha-helix or beta-sheet a simplified ‘ribbon diagram’ showing 2 polymer molecules of transthyretin that have deformed from random coils.

E N D
ENTROPYENTHALPY balances are the base of ALLpolymer self-organizing behaviour, such as the formation of an alpha-helix or beta-sheet a simplified ‘ribbon diagram’ showing 2 polymer molecules of transthyretin that have deformed from random coils
Even the most complex self-assembled structures of natural biopolymers (all proteins) (for example the beautifully intricate ‘beta-barrel’ structures of proteins such as streptavadin and porin) are all in essence very simple balances of coil ENTROPY and bond ENTHALPY
Our approach to predicting structure of proteins is based on an ENTROPY – ENTHALPY balance, where the coil ENTROPY wants to expand, But the folded structure is held in place by Enthalpic bonds such as: Hydrogen bonds, ionic bonds, hydrophobic bonds. (and we need only ΔG = ΔH — TΔS) to predict which structures will form at what T.
ΔG = ΔH — TΔS Tcritical = ΔH / ΔS most balances of ENTROPYENTHALPYare ΔH < 0 and ΔS < 0 randomassemble NO. crit. T YES ! cold hot
DNA is straightforward: a balance of only H-bonds and coil entropy. At a critical T, entropy wins out
Solving ΔG = ΔH — TΔSfor DNA with ΔH < 0 H-bonds balanced by ΔS < 0 deformation (enthalpy dominated state), defines a critical T (melting)
most balances of ENTROPYENTHALPYare ΔH < 0 and ΔS < 0 In self-assembled proteins the ENTHALPY wins over ENTROPY (H dominated) Therefore, an INCEASE in the temperature helps the ENTROPY term, and we observe random denaturing ΔG = ΔH — TΔS
Proteins are polypeptides prepared from 20 different amino acids, with a few different ΔH < 0 bonds holding them together, to form a vast number of folded structures which govern biological function and biomaterials.
In general, the natured state of proteins is ENTHALPY-dominated, and it requires much ENTHALPY to balance the ENTROPY to denature ΔS = -3/2[r2R0-2 + R02r-2] = ~40,000 kJ S penalty Q: how many H-bonds are needed to balance this? A: at ~50 kJ each, maybe about 800. (ignoring any hydrophobic bond help)
Most of the associative effects are ENTHALPIC in origin, except for the hydrophobic effect, which is essentially ENTROPIC in origin Here, it is the organized WATER molecules surrounding the polymer which increase their entropy by sharing a solvent cage
So the opposite effect occurs with balances where ENTROPY wins over ENTHALPY ΔG = ΔH — TΔS : (entropy dominated, eg: pyruvate carboxylase) Here, a DECEASE in the temperature lowers the ENTROPY term, and we observe denaturation. raise temp lower temp
The 3 basic ‘bonds’ holding proteins together each display a different strength dependence to their environment (temperature, pH, ions, ox/red chem.)
For example, IONIC bonds are controlled by pH (protonation changes charge), and the charge can also be screened by ions, changing both H and S
folding can be determined at 310 K for proteins knowing the allowed bond distances and angles, we can figure out how many H-bonds would be formed at all possible combinations of bond angles Psi and Phi :
a LOT of bonds get formed by wrapping the polymer around in a helix Either right-handed (easier) or left-handed (less easy) Thus, this structure occurs frequently.
This leads to a very common shape for proteins with set dimension (alpha helix) Abbreviated with a ‘coil’
The triple-wrap helix of collagen wraps again with many others, then again, and again until tendon, and connective tissue are formed. VERY strong.
Similar to the alpha helix, peptide chains lying side-by-side together match up for many bonds well, known as the beta sheet, either parallel or anti-parallel. Most proteins contain both elements.
but then it gets messy and very hard to predict… Hemoglobin full structure
One sample problem of protein adsorption (mostly to see how complex it can get in practice):The adsorption and spreading of a protein onto a surface (molecular recognition, rejection, attack): ΔspreadG = ΔadsorbH — TΔdeformS high coil entropy lower coil entropy but gain in enthalpy
h r the polymer chain is ‘N’ units of size ‘a’, and the equilibrium shape (with no H bonds) is Ro = aN3/5 the surface has ‘stickers’ of bond strength δ J/mol. what is radius r at eq ? exposed surface area under the coil = π r2 if the stickers are distance b apart, then the number of ‘stickers’ that this area covers is therefore = π r2 b–2
each bond has H gain –δ, ΔHtot = –δ π r2 b–2 J/mol initial coil volume = Ro3and the final volume V = hr2 if we consider the density to be conserved, we define h in terms of Ro & r: h = V r–2 = Ro3 r–2 (it is easiest to keep track of the coil height) the coil entropy ΔS = –3/2 [ h2/R02 + R02/h2 ] R ΔS = –3/2 [ r4/R04 + R04/r4 ] R
We have defined our ΔadsorbH and ΔdeformS , so : ΔspreadG = ΔadsorbH — TΔdeformS = –δ π r2 b–2 — –3/2 [ r4/R04 + R04/r4 ] RT ΔspreadG= –δ π r2 b–2 + 3/2 [ r4 R0–4 + R04 r–4 ] RT the units in both terms are J/mol. The equilibrium radius r can be determined as that which sets this equation to it’s minimum value.
ΔspreadG= –δ π r2 b–2 + 3/2 [ r4 R0–4 + R04 r–4 ] RT Q: When is this equation minimized ? A: when the first derivative w.r.t r is equal to 0. dG/dr = –2 δ π r b–2 + 6 RT [ r3 R0–4- R04r–5 ] dG/dr = 0 when : 6 RT r3 R0–4- 6 RT R04r–5 = 2 δ π r b–2
as a first test, we can see what happens as δ approaches zero (the absence of H bonding): dG/dr = 0 if: 6 RT r3 R0–4 – 6 RT R04r–5 = 2 δ π r b–2 with small δ : when 6 RT r3 R0–4 – 6 RT R04r–5 = 0 this means when 6 RT r3 R0–4 = 6 RT R04r–5 or when r8 = R08 meaning that in the absence of surface interactions, the coil maintains an isotropic conformation. With H-bonding however, the coil will deform, depending on the bond strength, the bond density (#bonds), and temperature
dG/dr = 0 if 3 RT r2 R0–4- 3 RT R04r–4 = δ π b–2 we are only concerned with the entropy term which describes the resistance to increasing r, so : dG/dr = 0 when 3 RT r2 R0–4 = δ π b–2 this means that r2 = (1 / 3 RT ) δ π b–2 R04 since Ro = aN3/5 r2 = (1 / 3 RT ) δ π b–2 a4 N12/5 if b = a (many bonds): r2 = (1 / 3 RT ) δ π a2 N12/5