Download

1 / 62
630 likes | 829 Views
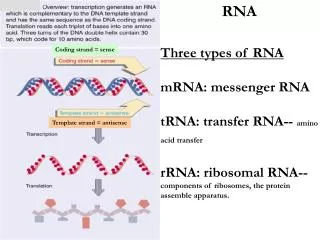
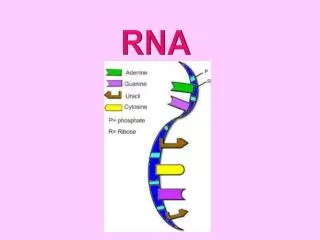
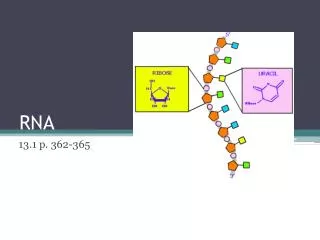
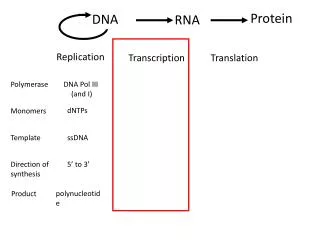
第五章 RNA 转录后的剪接与加工. 5.1 概述. 真核生物基因的表达在时间和空间上存在明显的 间隔,RNA转录在细胞核内进行,而翻译则在 细胞质内完成,因此不能像原核细胞那样几乎同 步同时地进行。. RNA processing. 5’ 端加帽 3’端形成 poly A 内含子的去除与外显子的连接 链的断裂 核苷酸修饰 糖苷键的改变 RNA 编辑. 5.2 原核生物 rRNA 的转录加工. rrn A- rrn G. m 4 Cm N 4 ,2’-O- 二甲基胞苷. 5.3 原核生物 tRNA 前体的加工.

E N D
第五章 RNA转录后的剪接与加工 5.1 概述 真核生物基因的表达在时间和空间上存在明显的 间隔,RNA转录在细胞核内进行,而翻译则在 细胞质内完成,因此不能像原核细胞那样几乎同 步同时地进行。
RNA processing • 5’端加帽 • 3’端形成poly A • 内含子的去除与外显子的连接 • 链的断裂 • 核苷酸修饰 • 糖苷键的改变 • RNA编辑
5.2 原核生物rRNA的转录加工 rrnA-rrnG
m4Cm N4,2’-O-二甲基胞苷
5.3 原核生物tRNA前体的加工 1. tRNA前体分子的形式: • 不同的tRNA串联排列 • 多个相同tRNA串联排列 • tRNA与rRNA混合串联排列
2.tRNA 前体的加工修饰 • 内切核酸酶使tRNA分子5’端成熟 • RNA外切酶使tRNA分子3’端成熟 • 在tRNA分子3’端添加CCAOH • 核苷酸的修饰与异构化
① tRNA分子3’端成熟 A.参与的酶: RNase P﹑ RNase F﹑ RNase D﹑ RNase BN﹑ RNase T﹑ RNase PH﹑ RNase Ⅱ﹑ polynucleotide PNPase
B. 步骤 • RNase P将tRNA前体分子水解,但是水解后的片段的3’端与5’端端仍还有额外的核苷酸 • RNase F从靠近3’端处水解切割,对tRNA前体3’端逐步加工 • RNase D从前体3’端再逐个切除附加序列,修剪tRNA的3’端 • 在tRNA核苷酰转移酶的作用下添加3’端CCA
② tRNA分子5’端成熟 由RNase P催化切除5’端额外核苷酸。该酶 是一种核糖核蛋白,由两个亚基组成,大亚 基是分子量约为125 kDa 的RNA,另外一个为 分子量为14 kDa的蛋白质。
5.5 真核生物RNA的转录后加工 (一)剪接方式的分类 ①第一类:自我剪接内含子,又可分为Ⅰ型和Ⅱ型内含子 ② 第二类:需蛋白质(酶)参与剪接的内含子 ③ 第三类:依赖sn RNP剪接的内含子
(二) 真核生物tRNA前体的转录后加工 1. 结构特点及加工概述 A. 与原核生物的差异 ①基因组内的tRNA基因成簇排列,各基因间有一定间隔, tRNA前体是单顺反子,且各个tRNA可作为独立的转录单位 ② tRNA基因数目比原核生物多 ③ tRNA基因有内含子,需经过剪接加工
B. 特点 • 长度和序列没有共同性,一般有16~46个核苷酸 • 位于反密码子的下游 • 内含子和外显子间的边界没有保守序列,因此tRNA前体的剪接方式不符合一般规律,需要RNase的参与 tRNA分子的二级结构:氨基酸受体臂、D环、TψC环和反密码环
2. 加工过程 • 内含子的剪接 tRNA前体分子中内含子的切除,两个tRNA半分 子的连接 • 3’端添加CCA:在tRNA核苷酸转移酶( tRNA nucleotide transferase)催化下进行3’端添加,由CTP和ATP提供胞苷酸和腺苷酸 • 核苷酸修饰 :甲基化酶、异戊烯转移酶、鸟嘌呤转糖苷酶、硫转移酶
(三) 真核生物rRNA前体的转录后加工 1. 结构组成与特点 • 小亚基: 16~18S rRNA • 大亚基: 26~28S rRNA 、 5S rRNA和5.8S rRNA • 基因拷贝数多,在基因组内成簇排列,成串重复数百次集中在核仁内,并由转录区(transcribed spacer)与非转录区(non-transcribed spacer, NTS)交替排列
2. rRNA前体加工的基本步骤 • 5’端非编码序列的切除, 41 S中间产物的生成 • 41S的RNA被切割成32 S(28S和5.8 S)与20S (18 S)两段 • 32 S被剪切成28S和5.8 S,并且部分序列互补配对 • 20 S被剪切成18 S 切割位点如何确定 ① 核仁小分子RNA (small nucleolar RNA, snoRNA)参与核糖核酸酶对特定立体结构的识别 ② rRNA前体分子的甲基化
3. snoRNA的结构与功能 ①结构特点 a. Box C框/D框,C框的序列为AUGAUGA, D框 为CUGA,可借助互补序列识别rRNA前体中甲基 化和切割的位点 b. Box H/ACA, H框为ANANNA,能识别假尿苷 酸化位点
② 功能 • 与蛋白质结合成snoRNP • 参与rRNA前体的加工 • box C/D具有互补序列,是指导rRNA中2’-O-核糖的甲基化修饰系统, box C参与甲基的转移反应 • box H/ACA能形成茎环二级结构,与rRNA特定序列互补
(四) 真核生物mRNA前体的加工 1. 5’端的帽子结构及加工步骤
核苷酸磷酸水解酶从生长着的RNA 5’端切去5’端γ磷酸基团 • 鸟苷酸7-甲基转移酶催化一分子游离GTP中的GMP攻击5’端第二个磷酸,形成5’,5’-三磷酸的连接,阻塞了RNA的5’端 • 2 ’-O-甲基转移酶催化S-腺苷甲硫氨酸(S-adenosyl methionine, AdoMet)的甲基基团转移到cap-0帽子结构鸟嘌呤的N7位 • 另外一分子甲酰基转移酶催化将另一分子的AdoMet连接到从5’端的第二个核苷酸cap-1的2-OH ’上,使之甲基化 • 继续由甲酰基转移酶催化随后的cap-2的2-OH ’甲基化及嘌呤核苷酸N7位甲基化,形成不同的帽子结构
2. 5’端帽子结构的功能 • 对mRNA的保护作用,是mRNA免受核苷酸酶从5’端开始的降解 • 使mRNA具有可翻译能力(translatability) • 有利于成熟mRNA从细胞核输送到细胞质
3. mRNA 3’端的多聚腺苷酸化(polyadenylation) • 保护mRNA,提高mRNA在细胞质中的稳定性 • 能增强mRNA的可翻译能力,在真核细胞的mRNA翻译过程中,有一种能结合在poly A上的蛋白质称为poly A结合蛋白I (poly A-bingding protein I, PAB I),当PAB I结合在poly A时,mRNA的翻译从效率大大增强
4.核内不均一RNA ①概述 核内不均一RNA(heterogeneous nuclear RNA, hnRNA)的碱基组成与总DNA组成类似,所以又 称为类似DNA的RNA(D-DNA) ② 与mRNA的关系 a.有相同的序列 b.在体外能作翻译模板,与mRNA有相同功能 c.两者的5’端都有帽子结构, 3’端有polyA d.两者的转录合成都可以被高剂量放线菌素D抑制
③hnRNA的结构特点 • 5’端有帽子结构(5’-cap) • 3’端有polyA+结构 • 帽子结构的下游3个寡聚U区,长度约30nt • 寡聚U区下游有重复序列 • 茎环结构分布于编码区的两侧 • 编码区为非重复结构,含有内含子
④hnRNA的加工过程 a. 5’端形成特殊的帽子结构(m7G5’ppp5’ N1mpN2p-) b. 修剪链的3’端,并加上polyA c. 通过剪接除去由内含子转录而来的序列 d. RNA链内的核苷酸被甲基化
5. mRNA前体加工的简要过程 ①mRNA前体中内含子的结构特点 a. 两端边界有共同序列,这些序列是mRNA前体 的剪接信号 b. 与第一类II型内含子剪接有些类似,必须形成 套索结构( lariat ),区别在于真核生物的mRNA的 剪接要求形成剪接体
②剪接体剪接的简要过程 a. 在内含子的5’切断,通过5’、 2’键将内含子5’- G与分支点A残基的2’为连接成套索结构 b. 在内含子的3’端剪接位点,外显子1与外显子2 连接,内含子以套索形式被释放
6. 真核生物mRNA前体的剪接机制 ①snRNP:由U1、U2、U4、U5 & U6 snRNP组成 a. U1 snRNP: 8中蛋白质与1种RNA链
b.U2 snRNP: 能与内含子的分支点共同序列互补,另外还与U6 snRNP碱基配对,协助snRNA在剪接体上定向
c.U4 snRNP:与U6 snRNP偶联形成配对的茎I和 茎II,当剪接体装配时,U4与U6解离开后发挥作 用。U4主要是结合U6直到在剪接需要U6时才释放 它,使之参与5’端剪接 d. U5 snRNP :与其它snRNA或剪接底物pre-mRNA 的保守区无明显的互补,但它的确同pre-mRNA的 5’和3’端剪接位点有相互作用
e. U6 snRNP 与内含子5’端剪接区域配对
②剪接体(spliceosome) a. 概念:mRNA前体在剪接过程中组装形成的多 组分复合物,主要是由细胞核内小分子RNA和蛋 白质组成 b. 组成:每种都含有1-2种snRNA和数种蛋白质因子
snRNP a.组成 SR蛋白 非snRNP SR相关蛋白:Tra、Tra2、U2AF ③剪接体的组装与循环
b. 步骤 • 早期剪接复合体的形成 • 前剪接复合体的组装 • 过渡复合物的形成 • 剪接体内结构重排 • 剪接产物的形成 • mRNA剪接产物和内含子的释放
Step 1. 早期剪接复合体的形成 • U1 snRNP结合到pre-mRNA的5’端剪接点上 • 剪接因子(splicing factor I)又称BBP (branch point binding protein)结合到分支点上 • U2 snRNP的辅助因子U2AF (U2 auxiliary factor)的65 kDa和35 kDa两个亚基分别结合到分支点和3’剪接位点的AG碱基上 • 在SR蛋白作用下,已经结合在5’剪接位点的U1snRNP和结合在3’剪接位点的SR/U2AF相互靠近,形成早期剪接复合物(early spliceosome complex)
Step 2. 前剪接复合体的组装 • 在U2辅助因子U2AF协助下,U2 snRNP与内含子的分支点结合 • ATP水解释放能量使内含子180°弯曲 • U1 snRNP和U2 snRNP两种复合物相互作用,使内含子5’端和3’端靠近 • SR蛋白协助U1 snRNP和U2AF结合,使早期剪接复合体组建成跨越内含子复合物,即前剪接体复合物(pre-spliceosome complex)
Step 3 过渡态复合物的形成 • U4/U6和U5 snRNP开始参与,使得U6内部序列和U4的5’端互补,前剪接体与U4 snRNP、U5 snRNP和U6 snRNP的三聚体结合 • U4与U6解离,U6在5’端剪接位点由ATP激活取代U1,使得U4离开前剪接复合体,同时在ATP的激活下,U1离开前剪接体复合物,形成过渡态复合物
Step 4 剪接体内的结构重排 • 在ATP激活下,U6与结合于分支点序列的U2 snRNP的5’端序列互补 • U1离开5’端剪接位点后,U5能够与两个外显子的剪接位点互补,使剪接体内发生结构重排,从而形成有利于催化剪接反应的构象
Step 5形成剪接产物并释放 • 在剪接体被激活的情况下,形成剪接产物 • 释放mRNA剪接产物和内含子
U5 snRNP环通过外显子1和外显子2接触,使两个剪接位点在 • 空间上靠近,由于它能引导其它RNA区域到剪接体的适当位置, • 以催化剪接功能的发挥,故又将U5 snRNP环称为导向RNA • b. U2 snRNP与内含子的分支点形成螺旋区域,分支点A周围的碱基 • 对使A突出,该结构为II型内含子的一种核酶(ribozyme)结构,是催 • 化剪接的活性中心
c. 剪接体组装过程中的重排反应 • 内含子5’端剪接点与U1 snRNP的相互作用转变为与U6 snRNP间的相互作用 • U2 snRNP与分支点相互作用代替了在分支点上结合蛋白BBP与分支点的相互作用 • U2 snRNP分子内部发生重排,形成茎-环构象的活化 • U4/U6之间RNA配对形成的茎II结构被破坏,U6 snRNP在其3’端形成分子内的茎-环结构 • U4/U6茎I区配对被破坏,使游离的U6 snRNP与U2 snRNP相互作用,而形成U2/U6螺旋I
U2 snRNP的5’茎-环结构被破坏使其5’端游离,并与U6相应部分作用形成U2/U6螺旋II 至此,前剪接体转变为成熟的剪接体,剪接催化 反应通过U4的释放启动,ATP水解供能
7. mRNA前体剪接中的转酯反应 • 概述:在成熟剪接体上完成剪接反应实际上是 磷酸二酯键的转移,又称为转酯反(transesterification) ①第一次转酯反应
②第二次转酯反应 • 第二次剪接反应是外显子1的3’端对内含子3’ 端剪接点进行亲核进攻,U5 snRNA在反应中起着 排布外显子的功能,使5’端外显子移动、定位,然 后在外显子I和II粗发生第二次转酯反应,形成5’, 3’-磷酸二酯键,产生两个外显子的连接物和一 个具有套索结构的内含子