Download

1 / 44
440 likes | 449 Views
This article discusses the synthesis and processing of glycans in the endoplasmic reticulum (ER) and Golgi apparatus, focusing on N-linked and O-linked oligosaccharides. It covers the biosynthesis of N-linked glycan precursors, the role of oligosaccharyltransferase complex (OST) in glycan transfer, and the initial processing of N-linked glycans. It also touches upon the role of ER processing in protein folding and quality control. Overall, it provides insights into the cellular context of glycan synthesis and processing.

E N D
Glycans in N-linkage to protein 1/23/07
Large O-linked Glycosaminoglycans and poly-lactosamine structures • Glycoprotein N-linked and O-linked oligosaccharides • Glycolipid oligosaccharides
Glycan synthesis in a cellular context
From ER to Trans-Golgi and points inbetween
“Hybrid” Major Classes of N-Glycans “Complex” “High-Mannose” (oligo-mannose) After Varki, A.
GlcNAc Man Glc Gal Sia Fuc Dolichol Biosynthesis of N-Glycans:Production of GlcNAc-P-P-Dolichol Tunicamycin Blocks - not very specific! Adapted from Marquardt T, Denecke J. Eur J Pediatr. 2003 Jun;162(6):359-79
GlcNAc Man Glc Gal Sia Fuc Biosynthesis of the N-GlycanPrecursor on the Cytosolic Leaflet of the Endoplasmic Reticulum (ER) CDG = Congenital Disorder of Glycosylation in Humans Adapted from Marquardt T, Denecke J. Eur J Pediatr. 2003 Jun;162(6):359-79
GlcNAc Man Glc Gal Sia Fuc Biosynthesis of the N-GlycanPrecursor on Lumenal Leaflet of ER Adapted from Marquardt T, Denecke J. Eur J Pediatr. 2003 Jun;162(6):359-79
GlcNAc Man Glc Gal Sia Fuc Completion of Biosynthesis of N-GlycanPrecursor on Lumenal Leaflet of ER- and Transfer to Protein Adapted from Marquardt T, Denecke J. Eur J Pediatr. 2003 Jun;162(6):359-79
Target “sequon” for N-glycosylation • Necessary but not sufficient • X = any amino acid except proline • Rarely can be Asn-X-Cys • Transfer co-translational/immediate • post-translational before folding • ~2/3 of proteins have sequons • ~ 2/3 sequons actually occupied • (some variably) Yeast OST complex contains nine membrane-bound subunits Oligosaccharyltransferase complex (OST) in the ER membrane transfers the dolichol N-glycan precursor to asparagine residues on nascently translated proteins
GlcNAc Man Glc Gal Sia Fuc Initial Processing of N-Glycans in the ER and Golgi ER Golgi Adapted from Marquardt T, Denecke J. Eur J Pediatr. 2003 Jun;162(6):359-79
Improperly folded proteins are re-glucosylated by a glucosyltransferase which acts as “sensor” for improper folding Deglucosylation/glucosylation cycles facilitate protein folding, thereby controling the quality of the glycoproteins produced in the ER Jones, J., 2005, BBA 1726, 121-137
Processing enzymes in the ER prepare the Glc3Man9GlcNAc2 precursor for subsequent processing in the Golgi Jones, J., 2005, BBA 1726, 121-137
General Principles for ER Processing of N-linked glycans • Precursors begin as lipid-linked species on the cytoplasmic face of the ER, requiring that substrates be flipped for further processsing • Donor substrates on the cytoplasmic face of the ER membrane are nucleotide sugars • Donor substrates on the lumenal face of the ER are dolichol-P-sugars • Addition of N-linked glycans occurs co-translationally through the activity of a multi-protein complex • ER glycosylation is well-conserved and most likely evolved as a method for quality control • Degradative steps in ER-glycan processing are coupled to checkpoints that ensure the export of functional protein from the ER
N-acetylglucosamine Glucose Sialic acid Mannose Galactose Fucose N-linked Oligosaccharide Processing Pathway “high mannose” ER Lumen Cytosol Trimming and processing Synthesis of lipid- linked precursor Glycan transfer “complex” P P Golgi Lumen Cytosol Terminal glycosylation
“hybrid” GlcNAc Sialic acid Glucose Mannose Fucose Galactose “high mannose” newly synthesized N-linked glycan “complex”
slime mold yeast Xylose GlcNAc Sialic acid Glucose Mannose Fucose Galactose Evolutionary diversity in N-glycans conserved in fungi, plant and mammals vertebrates plants Drosophila C. elegans
N-acetylglucosamine Glucose Sialic acid Mannose Galactose Fucose N-linked Oligosaccharide Processing Pathway “high mannose” ER Lumen Cytosol Trimming and processing Synthesis of lipid- linked precursor Glycan transfer “complex” P P Golgi Lumen Cytosol Terminal glycosylation
P P Man GlcNAc Phosphorylation of mannose residues is specific to lysosomal enzyme oligosaccharides and occurs via a two-step process P- -P UDP- UMP GlcNAc-1- Phosphotransferase “Uncovering” enzyme Phosphotransferase:alpha/beta subunits - catalytic activity gamma subunit - specific recognition of lysosomal enzymes
Lysosomal Enzymes Bearing Man-6-P Residues are Sorted to Lysosomes by Two Mannose-6-phosphate Receptors (CD-MPR: 46 kDa and CI-MPR: 300 kDa) secreted glycoprotein lysosomal enzyme MPR
Lysosomal enzymes lacking mannose-6-phosphate on their N-linked oligosaccharides are secreted from the cell (genetic deficiency in GlcNAc-1-phosphotransferase)
fibroblasts from a patient with mucolipidosis II (I-cell disease)
N-acetylglucosamine Glucose Sialic acid Mannose Galactose Fucose N-linked Oligosaccharide Processing Pathway “high mannose” ER Lumen Cytosol Trimming and processing Synthesis of lipid- linked precursor Glycan transfer “complex” P P Golgi Lumen Cytosol Terminal glycosylation
GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan synthesis: post-ER mannose trimming Golgi 1,2-mannosidase I Man8-9 Man5 • three paralogs (IA, IB, IC) have essentially identical enzymatic specificity but • display distinct developmental and tissue-specific expression patterns
ManII GnT-I GnT-II Golgi mannosidase II ß1,2 GlcNAcT-I ß1,2 GlcNAcT-II Endo H-resistant Endo H-sensitive Endo H treatment is a useful measure of a glycoprotein’s arrival in the Golgi GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan synthesis: initiation of branch or antennae formation N-glycans can exhibit different sensitivity to bacterial glycosidases (endo-ß-N-acetylglucosaminidase H or Endo H)
ManII GnT-I GnT-II Golgi mannosidase II ß1,2 GlcNAcT-I ß1,2 GlcNAcT-II mouse knockout of Man-II: dyserythropoietic amenia Why? GlcNAc Sialic acid Glucose Mannose Fucose Galactose The biosynthesis of complex type N-glycans is essential for development mouse knockout of GnT-I: embryonic lethal mouse knockout of GnT-II: early post-natal lethality
ManII GnT-I GnT-II Golgi mannosidase II ß1,2 GlcNAcT-I ß1,2 GlcNAcT-II MX Golgi mannosidase IIx Golgi mannosidase IIx is not expressed in hematopoietic cells (no bypass pathway ---> dyserythropoiesis ---> anemia) ManII and MX double null mice exhibit pre- and postnatal lethality and complete absence of complex type N-glycans
+ endomannosidase glucosidase inhibitors (castanospermine), deficiency of glucosidase enzymes Other examples of bypass mechanisms: endomannosidase ER glucosidase II ER glucosidase I complex
GlcNAcT-III ß4 GlcNAcT-II ß2 GlcNAcT-I ß2 GlcNAcT-V ß6 GlcNAcT-IV ß4 GlcNAcT-VI ß4 GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan synthesis: branching via GlcNAc addition branch = antennae
GlcNAcT-V (GnT-V) - adds the ß1,6 branch to complex type N-glycans • upregulation of GnT-V (and increased formation of ß1,6 branch) has been associated • with increased tumorigenesis and metastasis • increased ß1,6 branch formation of specific glycoproteins might alter their function • (e.g. integrin, proteases) • GnT-III expression (increased bisecting GlcNAc addition) appears to antagonize the • effects of GnT-V (suppression of lung metastasis by melanoma cells when GnT-III • is introduced) • - upregulation of GnT-III is also tumorigenic in some cases
GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-glycan diversity is generated through branch modification developmentally-regulated tissue-specific protein-specific
GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan synthesis: galactosylation ßGalT1 ß1,4-galactosyltransferase • ß1,4GalTs utilize many acceptors (including GlcNAc-terminating branches of N-glycans) • for the addition of galactose • seven ß1,4GalT genes in mammalian genomes • ß1,4GalT has two enzyme activities: it transfers Gal to GlcNAc-based acceptors and • when associated with -lactalbumin, it transfers Gal to Glu to form lactose (ß1,4GalT • expression is induced in mammary glands during lactation)
2,3-sialyltransferase 2,6-sialyltransferase GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan branch modification: sialylation SiaTs STX, PST Poly 2,8-sialyltransferase • changes in sialylation are associated with tumor metastatic potential • sialic acids play important roles in the immune system • polysialic acid is predominantly found on the neural cell adhesion molecule (NCAM)
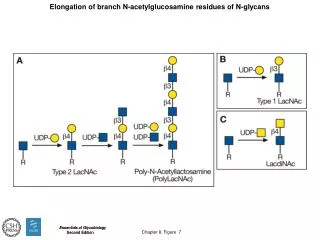
GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan branch modification: polylactosamine biosynthesis iGnT, ßGalTI ß1,3 GlcNAcT, ß1,4 GalTI • found on many red blood cell glycoproteins (functions not clear) • preferred substrate for galectin-1
GalNAc GlcNAc N-linked glycan branch modification: diversity in terminal modifications (sulfation, GalNAc addition, fucosylation, etc.) found on the oligosaccharides on pituitary hormones, LH and TSH (but not FSH) Some of these structures are unique to N-glycans but many can also be found on O-glycans and glycolipids
GlcNAc Sialic acid Glucose Mannose Fucose Galactose N-linked glycan synthesis: core fucosylation Fut8 1,6-fucosyltransferase • levels of core fucosylation altered in certain cancers but no functional • insight until Fut8 -/- mice generated Wang X. et. Al. PNAS (2005) vol. 102, p: 15791-15796
lung tissue sections surviving mice exhibit emphysema-like changes in lung
What proteins are associated with this lung pathology? Overexpression of MMPs and down-regulation of ECM proteins Deficiency in TGFß1 signaling Binding of 125I-TGFß1 is reduced, SMAD2 phosphorylation and TGFß receptors are underfucosylated in Fut8 -/- mouse fibroblasts Wild type Fut8 KO Fut8 KO + Fut8 AAL: lectin that binds to core fucose
surviving postnatal 18-day-old Fut8-/- mice were treated with or without recombinant TGF-1 Lung tissue sections Conclusion: Fut8 -/- TGFß1 receptor is partially deactivated by lack of core fucose and therefore excess TGFß1 can overcome the block
Elucidating the function of N-glycans • cultured cells that lack the expression of specific enzymes involved • in N-glycan biosynthesis • mouse knockouts of glycosyltransferases • experiments in nature (CDGs) CDGs: Congenital Disorders of Glycosylation • a group of clinically heterogenous autosomal recessive human disorders that • are characterized by mutations in genes that encode glycosylation enzymes • defects are leaky (hypomorphic alleles)
CDGs: defects in oligosaccharide biosynthesis and processing ER CDG-I c-h,l Lumen CDG-IIb Cytosol CDG-I a,b,i,j,k Golgi P P MLII, MLIII CDG-IIa CDG-IId CDG-II c Glucose N-acetylglucosamine Sialic acid GDP- Mannose Fucose Galactose CDG-IIf CMP-
N-linked glycosylation: the necessity for both consistency and diversity • the biosynthesis of the 14-sugar lipid-linked oligosaccharide precursor has little • room for error • Why is there so much diversity in the final product? Gagneux and Varki “Evolutionary considerations in relating oligosaccharide diversity to biological function” Glycobiology (2006) p: 747-755
Key points: • N-linked oligosaccharides can be found in three main types in mammalian tissues: • HIGH MANNOSE, COMPLEX and HYBRID • The structural diversity of N-glycans in many organisms (e.g. invertebrates) has been • underestimated (more complex structures are detected but very rare) • The addition of mannose 6-phosphate residues onto the high mannose type • oligosaccharides of lysosomal enzymes mediates their targeting to lysosomes • Coordinated action of a glycosidase (Man-II) and two glycosyltransferases (GnT-I and • GnT-II) initiate the biosynthesis of complex type N-glycans • Degree of N-glycan branching is dictated by the addition of GlcNAc residues (2-4 • branches or antennae most common) • Extensive structural diversity is present in the terminal modification of N-glycans; • these terminal modifications often control glycan function • Formation of complex type N-glycans is essential for development (so important • that several mechanisms to avoid blocks in N-glycan processing have evolved)