Download

1 / 49
963 likes | 2.13k Views
Mechanisms of Antibiotic Resistance. What is antibiotic resistance?.

E N D
What is antibiotic resistance? • Antibiotic resistance occurs when an antibiotic has lost its ability to effectively control or kill bacterial growth; in other words, the bacteria are "resistant" and continue to multiply in the presence of therapeutic levels of an antibiotic.
Why do bacteria become resistant to antibiotics? • When antibiotics are used to kill the bacterial microorganisms, a few microorganisms are able to still survive, because microbes are always mutating, eventually leading to a mutation protecting itself against the antibiotic
Antibiotics that are used correctly overwhelm the harmful bacteria • Overuse of antibiotics or unnecessary use creates a selective environment • Resistant bacteria has better fitness in this context • Resistant strains survive and multiply. • After reproducing, the resistant bacteria move to another host.
Sex in bacteria • Bacteria do exchange genes forming new combinations • Bacteria exchange genes is by conjugation • This involves the transfer of genetic material via a cytoplasmic bridge between the two organisms • This can be done between unrelated species of bacteria • Recent studies on bacteria in the wild show that it definitely occurs in the soil, in freshwater and oceans and inside living organisms
The magic bullet • Antibiotics revolutionised medicine • The first antibiotic, penicillin, was discovered by Alexander Fleming in 1929 • It was later isolated by Florey and Chain • It was not extensively used until the 2nd World War when it was used to treat war wounds • After 2nd World War many more antibiotics were developed • Today about 150 types are used • Most are inhibitors of the protein synthesis, blocking the 70S ribosome, which is characteristic of prokaryotes
Resistance • It took less than 20 years for, bacteria to show signs of resistance • Staphylococcus aureus, which causes blood poisoning and pneumonia, started to show resistance in the 1950s • Today there are different strains of S. aureus resistant to every form of antibiotic in use
Multiple resistance • It seems that some resistance was already naturally present in bacterial populations • The presence of antibiotics in their environment in higher concentrations increased the pressure by natural selection • Resistant bacteria that survived, rapidly multiplied • They passed their resistant genes on to other bacteria (both disease causing pathogens and non-pathogens)
Transposons & Integrons • Resistance genes are often associated with transposons, genes that easily move from one bacterium to another • Many bacteria also possess integrons, pieces of DNA that accumulate new genes • Gradually a strain of a bacterium can build up a whole range of resistance genes • This is multiple resistance • These may then be passed on in a group to other strains or other species
Antibiotics promote resistance • If a patient taking a course of antibiotic treatment does not complete it • Or forgets to take the doses regularly, • Then resistant strains get a chance to build up • The antibiotics also kill innocent bystanders bacteria which are non-pathogens • This reduces the competition for the resistant pathogens • The use of antibiotics also promotes antibiotic resistance in non-pathogens too • These non-pathogens may later pass their resistance genes on to pathogens
How humans have created the upsurge of bacterial diseases: • International travel • Inadequate sanitation • “antibiotic paradox”
Antibiotic Resistance – A Global Problem VRE PRP VISA ESBL MBL MRSA VRSA 1996 2002 1986 1961 1967 1983 1988 Vancomycin and teicoplanin Penicillin Vancomycin Vancomycin and teicoplanin 3rd gencephalosporin All -lactams Carbapenem Emergence→Spread
How do bacteria become resistant? Bacteria can gain resistance over time through: • Acquired resistance • Vertical gene transfer • Horizontal gene transfer
Genetic Basis of Resistance • Spontaneous mutations in endogenous genes • Structural genes: expanded spectrum of enzymatic activity, target-site modification, transport defect • Regulatory genes: increased expression • Acquisition of exogenous genes • Usually genes that encode inactivating enzymes or modified targets, regulatory genes • Mechanisms of DNA transfer: conjugation (cell–cell contact); transformation (uptake of DNA in solution); transduction (transfer of DNA in bacteriophages) • Expression of resistance genes • Reversible induction/repression systems can affect resistance phenotypes
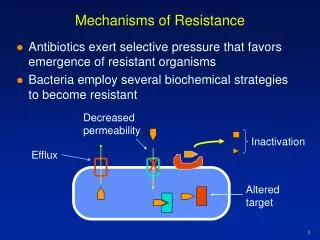
Mechanisms of Resistance • Antibiotics exert selective pressure that favours emergence of resistant organisms • Bacteria employ several biochemical strategies to become resistant
Major Classes of Antibiotics PBPs penicillin-binding proteins
b-Lactams: Classification (1) • Penicillins • Narrow-spectrum penicillins • Broad-spectrum penicillins • β-lactamase inhibitor combinations • Oxacillin derivatives • Cephalosporins (ATC/WHO 2005 classification) • 1st generation: Gram-positive cocci (GPCs), some Gram-negative bacilli (GNBs) • 2nd generation: some GNBs, anaerobes • 3rd generation: many GNBs, GPCs • 4th generation: many GNBs resistant to 3rd generation, GPCs
b-Lactams: Classification (2) • Carbapenems • Imipenem, meropenem, Doripenem, ertapenem • Monobactams • Aztreonam
Cytoplasm NAG-NAM-NAG-NAM | L-Ala | D-Glu | L-diA | D-Ala | D-Ala NAG-NAM-NAG-NAM | L-Ala | D-Glu | L-diA | D-Ala | D-Ala -(AA)n-NH2 -(AA)n-NH2 Transpeptidation reaction Mechanism of Action of b-Lactams (1) • Structure of peptidoglycan
Mechanism of Action of b-Lactams (2) • Penicillin-binding proteins (PBPs) • Membrane-bound enzymes • Catalyse final steps of peptidoglycan synthesis (transglycosylation and transpeptidation) • -lactams • Act on PBPs, inhibit transpeptidation • Substrate analogues of D-Ala-D-Ala
Resistance to b-Lactams • Gram-negative b-lactamases • Major resistance mechanism in nosocomial GNB pathogens • >470 b-lactamases known to date • Classified into 4 groups based on sequence similarity • Ambler Class A (TEM, SHV, CTX), C and D (OXA) are serine b-lactamases • Ambler Class B are metallo-b-lactamases • Their spread has been greatly exacerbated by their integration within mobile genetic elements • Integron-borne b-lactamase genes are part of multi drug resistance gene cassettes Multidrug-resistant nosocomial pathogens with complex resistance patterns Selection of potent b-lactamases through use of non-b-lactam agents
Ambler Classification of β-Lactamases β-lactamases Active site Serine-enzymes Zinc-enzymes Nucleotide sequence C D B A Four evolutionarily distinct molecular classes
Modified Bush–Jacoby–Medeiros Classification of b–Lactamases
b-Lactamases: Classification Serine enzymes Metallo (Zn) enzymes Group C Group A Group D Group B AmpC TEM/SHV OXA IMP/VIM Cephs Pens, Cephs Pens, esp Oxa Carbapenems Inhib-R Inhib-S Inhib-R/S Inhib-R Bush. Rev Inf Dis 1987;10:681; Bush et al. Antimicrob Agents Chemother 1995;39:12; Bush. Curr Opin Investig Drugs 2002;3:1284
Induction of Group 1 (AmpC) b-Lactamases Inducible: Enterobacter spp. Citrobacter spp. Morganella spp. Providencia spp. Serratia spp. P. aeruginosa Amount enzyme per cell Absent : Salmonella spp. Klebsiella spp. Basal : E. coli Shigella spp. b-lactam concentration
Selection of Group 1 (AmpC)b-Lactamases Population of inducible organisms Derepressed cell due to ampD mutation (Enterobacter : 1 of 105 !) Ceftazidime, ceftriaxone, piperacillin, etc.: Selection of derepressed cell Multiplication and spread of derepressed clone
Group 1 (AmpC) b-Lactamases • Produced constitutively in tiny concentrations by certain GNB • Induction of production: • Can occur by the exposure to certain antibiotics (eg, carbapenems) • Only in vitro phenomenon; not clinically relevant (stops when antibiotic use is discontinued; carbapenems not affected by these enzymes) • Selection of production: • Can occur by the use of certain antibiotics (eg, ceftazidime) • Also in vivo phenomenon; highly clinically relevant (does not stop when antibiotic use is discontinued; leads to selection and spread of ABR clones) • Therapeutic options: • 4th generation cephalosporins (but resistance may occur with minor AA changes) • Carbapenems
Resistance to b-Lactams • Chromosomal AmpCb-lactamases • Several Enterobacteriaceae, including Enterobacter, Citrobacter, and Serratia contain an inducible, chromosomal gene coding for a b-lactamase • Resistant to cephalosporins and monobactams; not inhibited by clavulanate; Class C b-lactamases • Plasmid-mediated AmpCb-lactamases • Arose through transfer of AmpC chromosomal genes into plasmids • Not inducible, with substrate profile (usually) same as parental enzyme • Highly prevalent in the naturally AmpC-deficient K. pneumoniae • Emergence predominantly in community-acquired infections (Salmonella spp., E. coli) • Co-resistance to aminoglycosides, SXT, quinolones • Wide dissemination worldwide (SE Asia, N Africa, South Europe, USA)
Plasmid-mediated AmpCb-lactamases (1) Enzyme Host Country Year isolated MIR-1 K. pneumoniae US 1988 ACT-1 K. pneumoniae US 1994 E. coli BIL-1 E. coli UK 1989 CMY-2 K. pneumoniae Greece 1990 S. senftenberg France 1994 Salmonella US 1996 E. coli Libya 1996 Salmonella Spain 1999 Salmonella Romania 2000 LAT-1 K. pneumoniae Greece 1993 LAT-2 K. pneumoniae, Greece 1994 E. coli, E. aerogenes CMY-3 P. mirabilis France 1998 CMY-4 P. mirabilis Tunisia 1996 E. coli UK 1999 K. pneumoniae Sweden 1998 CMY-5 K. oxytoca Sweden 1988 CMY-7 E. coli India
Plasmid-mediated AmpCb-lactamases (2) Enzyme Host Country Year isolated DHA-1 Salm. enteritidis Saudi Arabia 1992 K. pneumoniae Taiwan 1999 US 1996-2000 DHA-2 K. pneumoniae France 1992 ACC-1 K. pneumoniae Germany 1997 K. pneumoniae France 1998 P. mirabilis Tunisia 1997 K. pneumoniae Tunisia 1999 Salm. livingstone Tunisia 2000
Plasmid-mediated AmpCb-lactamases (3) Enzyme Host CountryYear isolated FOX-1 K. pneumoniae Argentina 1989 FOX-2 E. coli Germany 1993 FOX-3 K. oxytoca, Italy 1994 K. pneumoniae FOX-4 E. coli Canaries 1998 FOX-5 K. pneumoniae US 1999 CMY-1 K. pneumoniae Korea 1989 CMY-8 K. pneumoniae Taiwan 1998 CMY-9 E. coli Japan 1995 CMY-10 E. aerogenes Korea 1999 CMY-11 E. coli Korea 1998 MOX-1 K. pneumoniae Japan 1991 MOX-2 K. pneumoniae France 1999
Resistance to b-Lactams • Extended-spectrum b-lactamases (ESBL) • No consensus of the precise definition of ESBLs • In general: β-lactamases conferring resistance to the penicillins, 1st , 2nd, 3rd, and even 4th generation cephalosporins, and monobactams, not to carbapenems and cephamycins • Inhibited by b-lactamase inhibitor clavulanic acid • Derived from Class A b-lactamases (exceptions are Class D, OXA): TEM, SHV, CTX-M, OXA, VEB, PER,... • Differ from their progenitors by 1–5 amino acids • Marked and unexplained predilection for Klebsiellapneumoniae • Therapeutic options: carbapenems
The Story of E. coli Resistant to Ampicillin • June 1964: ampicillin released in Europe • December 1964; the first case of ampicillin- resistant E. coli detected • Mrs Temoneira (Athens, Greece): • Urinary isolate of E. coli • Produced b-lactamase (TEM-1) • Genes encoding the b-lactamase found on a plasmid
The Story of Klebsiella UniversallyResistant to Ampicillin • SHV-1 enzyme: b-lactamase with a narrow spectrum of activity (ampicillin) • Chromosomally encoded • If produced in high amounts: • May result in resistance to cefazolin and piperacillin • May even overcome β-lactamase inhibitors (clavulanic acid or tazobactam)
The Story of ESBL-producingEnteric GNB • Third generation cephalosporins: • Developed in response to proliferation of K. pneumoniae and E. coli producing b-lactamases active against ampicillin and first generation cephalosporins • Introduced in Europe in the early 1980s • Emergence of extended-spectrum b-lactamases: • Cefotaxime marketed in Germany in September 1981 • Cefotaxime-resistant Klebsiella isolate detected in Frankfurt in March 1982 (mutant of the gene encoding SHV-1)
MIC (mg/mL) ceftazidime 102 162 TEM-1 0.25 glutamine arginine TEM-12 2.0 glutamine serine TEM-26 128 lysine serine Evolution of TEM Enzymes
ESBLs in Non-fermenters • Emergence of transferable ESBL enzymes (Class A, B or D) in non-fermenters (P. aeruginosa, Acinetobacter spp.) • ESBL types often different (PER-1, VEB-1, OXA,…) from Enterobacteriaceae • Multiple resistance mechanisms co-expressed (chromosomal AmpC b-lactamase, impermeability, efflux) • Non-fermenters should not be tested routinely for ESBLs • P. aeruginosa: «False-negative» (most ESBLs not inhibited by clavulanate) • Acinetobacter spp.: «False-positive» DD with clavulanate (intrinsic activity of b-lactam inhibitors) • S. maltophilia: «False-positive» DD with clavulanate (inhibition of L2 chromosomal enzyme)
Current ESBL Detection Methods Fail • Routine tests are not designed for ESBL detection • Low level ESBL expression will not be detected by current tests using low inoculum • MIC values and zone sizes of ESBL producers overlap those of susceptible non-ESBL producers • ESBL double disk test may be inaccurate if positioning is suboptimal • ESBL breakpoint methods are limited since MICs for different strains can range over 7 dilutions
Resistance to b-Lactams • Carbapenemases • Defined as b-lactamases, hydrolyzing at least imipenem or/and meropenem or/and ertapenem • Belong to Ambler Class A, B, and D, of which Class B are the most clinically significant: • Class A: KPC, SME & NMC/IMI • Class B: IMP, VIM & SPM metallo b-lactamases • Class D: OXA-23, -40 & -58 related
Class B (Metallo)-Carbapenemases • Hydrolyzing virtually all b-lactams • Mediate broad spectrum b-lactam resistance • No clinical inhibitor available • Present on large plasmids and integrons • Genes are continuously spreading • Associated (80%) with aminoglycoside resistance Still rare but increasing, especially in non-fermenters
Mobile Carbapenamases Class I integron • Nosocomial outbreak of carbapenem-resistant P.aeruginosa and A. baumanii reported in Canada and France, respectively • Cross-resistance to other beta-lactams and to other AB classes • Link with aminoglycoside use, not necessarily carbapenems! blaIMP blaVIM ORF1 aacC4 aacC1 5'cs 3'cs
Mobile Class B b-Lactamases Enzyme Host Country (Year) IMP-1S. marcescensJapan (>91) P. aeruginosaJapan A. xylosoxydansJapan P. putidaJapan C. freundiiJapan K. pneumoniaeJapan, Singapore (99) A. baumanniiJapan P. stutzeri, Taiwan P. putida A. junii UK (00) IMP-2A. baumanniiItaly (97) IMP-3 S. flexneriJapan (96) IMP-4 Acinetobacter Hong Kong (>94) C. youngae China (98) IMP-5A. baumannii Portugal (98) IMP-6S. marcescensJapan (96) IMP-7P. aeruginosaCanada (95) Malaysia (99) IMP-8K. pneumoniae Taiwan (98) IMP-9P. aeruginosa China (?) IMP-10A. xylosoxydansJapan (00) P. aeruginosaJapan (97) Enzyme Host Country (Year) VIM-1P.aeruginosaItaly (1997) A. baumanniiItaly (1997) P.aeruginosaGreece (1996) E. coliGreece (2001) A. xylosoxydansItaly (1997) VIM-2P. aeruginosaFrance (1996) P. aeruginosaGreece (1996) P. aeruginosaItaly (1998) S. marcescensKorea (2000) A. baumanniiKorea (1998) P. aeruginosaBelgium (2004/5) P.putidastutzeri Taiwan (>1997) VIM-3P. aeruginosa Taiwan (>1997) VIM-4P. aeruginosaGreece (2001) SPM-1P. aeruginosa Brazil (1997) GIM-1P. aeruginosa Germany (2003)
Class D Oxacillinase — Carbapenemases • Class D enzymes • OXA-23, -24, -25, -26, -27, -28, -40, -49, -58, …. • Highly mobile (integron, plasmid) • Found in South America, South-East Asia, Europe (Greece, Spain, Portugal, France, Belgium) • Multi-drug resistance (penicillins and 3rd & 4th generation cephalosporins, BL/BL-inhibitors, aminoglycosides, SXT,…) • Variable resistance levels to imipenem and meropenem (4–>256 mg/mL)
Rapidly Increasing Antibiotic Resistance Constitutes One of the Most Important Clinical, Epidemiological and Microbiological Problems of Today