Download
1 / 1
10 likes | 353 Views
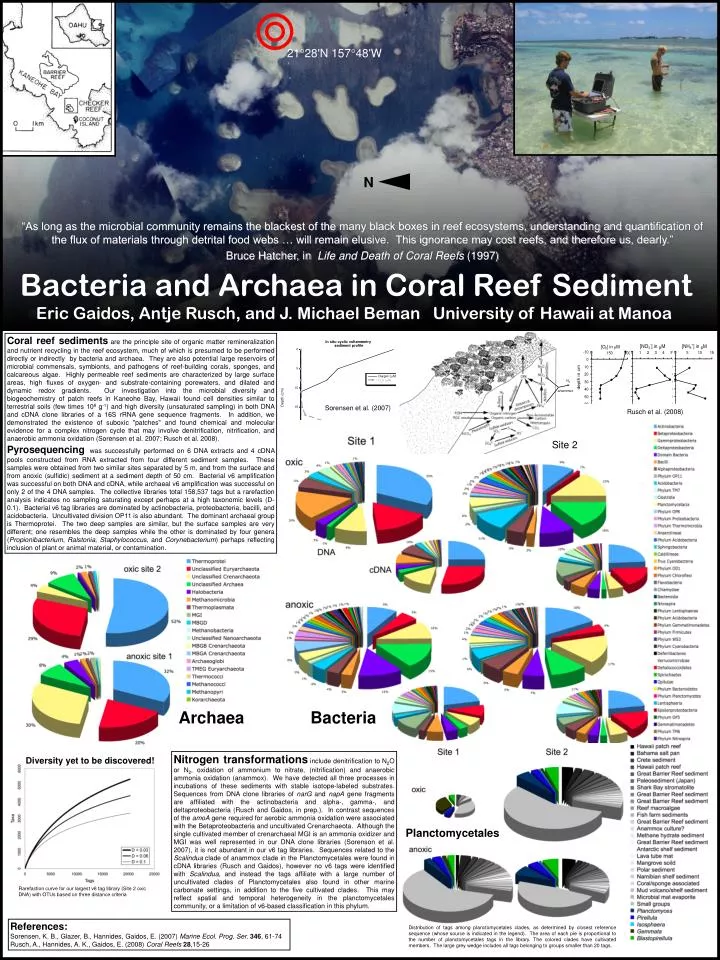
21 28'N 157 48'W. N. “As long as the microbial community remains the blackest of the many black boxes in reef ecosystems, understanding and quantification of the flux of materials through detrital food webs … will remain elusive. This ignorance may cost reefs, and therefore us, dearly.”

E N D
2128'N 15748'W N “As long as the microbial community remains the blackest of the many black boxes in reef ecosystems, understanding and quantification of the flux of materials through detrital food webs … will remain elusive. This ignorance may cost reefs, and therefore us, dearly.” Bruce Hatcher, in Life and Death of Coral Reefs (1997) Bacteria and Archaea in Coral Reef Sediment Eric Gaidos, Antje Rusch, and J. Michael Beman University of Hawaii at Manoa Coral reef sediments are the principle site of organic matter remineralization and nutrient recycling in the reef ecosystem, much of which is presumed to be performed directly or indirectly by bacteria and archaea. They are also potential large reservoirs of microbial commensals, symbionts, and pathogens of reef-building corals, sponges, and calcareous algae. Highly permeable reef sediments are characterized by large surface areas, high fluxes of oxygen- and substrate-containing porewaters, and dilated and dynamic redox gradients. Our investigation into the microbial diversity and biogeochemistry of patch reefs in Kaneohe Bay, Hawaii found cell densities similar to terrestrial soils (few times 108 g-1) and high diversity (unsaturated sampling) in both DNA and cDNA clone libraries of a 16S rRNA gene sequence fragments. In addition, we demonstrated the existence of suboxic "patches" and found chemical and molecular evidence for a complex nitrogen cycle that may involve denitrification, nitrification, and anaerobic ammonia oxidation (Sorensen et al. 2007; Rusch et al. 2008). Pyrosequencingwas successfully performed on 6 DNA extracts and 4 cDNA pools constructed from RNA extracted from four different sediment samples. These samples were obtained from two similar sites separated by 5 m, and from the surface and from anoxic (sulfidic) sediment at a sediment depth of 50 cm. Bacterial v6 amplification was successful on both DNA and cDNA, while archaeal v6 amplification was successful on only 2 of the 4 DNA samples. The collective libraries total 158,537 tags but a rarefaction analysis indicates no sampling saturating except perhaps at a high taxonomic levels (D-0.1). Bacterial v6 tag libraries are dominated by actinobacteria, proteobacteria, bacilli, and acidobacteria. Uncultivated division OP11 is also abundant. The dominant archaeal group is Thermoprotei. The two deep samples are similar, but the surface samples are very different; one resembles the deep samples while the other is dominated by four genera (Propionibacterium, Ralstonia, Staphylococcus, and Corynebacterium) perhaps reflecting inclusion of plant or animal material, or contamination. N2 anammox Sorensen et al. (2007) Rusch et al. (2008) Site 2 Archaea Bacteria Nitrogen transformationsinclude denitrification to N2O or N2, oxidation of ammonium to nitrate, (nitrification) and anaerobic ammonia oxidation (anammox). We have detected all three processes in incubations of these sediments with stable isotope-labeled substrates. Sequences from DNA clone libraries of narG and napA gene fragments are affiliated with the actinobacteria and alpha-, gamma-, and deltaproteobacteria (Rusch and Gaidos, in prep.). In contrast sequences of the amoA gene required for aerobic ammonia oxidation were associated with the Betaproteobacteria and uncultivated Crenarchaeota. Although the single cultivated member of crenarchaeal MGI is an ammonia oxidizer and MGI was well represented in our DNA clone libraries (Sorenson et al. 2007), it is not abundant in our v6 tag libraries. Sequences related to the Scalindua clade of anammox clade in the Planctomycetales were found in cDNA libraries (Rusch and Gaidos), however no v6 tags were identified with Scalindua, and instead the tags affiliate with a large number of uncultivated clades of Planctomycetales also found in other marine carbonate settings, in addition to the five cultivated clades. This may reflect spatial and temporal heterogeneity in the planctomycetales community, or a limitation of v6-based classification in this phylum. Diversity yet to be discovered! Planctomycetales Rarefaction curve for our largest v6 tag library (Site 2 oxic DNA) with OTUs based on three distance criteria References: Sorensen, K. B., Glazer, B., Hannides, Gaidos, E. (2007) Marine Ecol. Prog. Ser.346, 61-74 Rusch, A., Hannides, A. K., Gaidos, E. (2008) Coral Reefs28,15-26 Distribution of tags among planctomycetales clades, as determined by closest reference sequence (whose source is indicated in the legend). The area of each pie is proportional to the number of planctomycetales tags in the library. The colored clades have cultivated members. The large grey wedge includes all tags belonging to groups smaller than 20 tags.