Download

1 / 51
510 likes | 676 Views
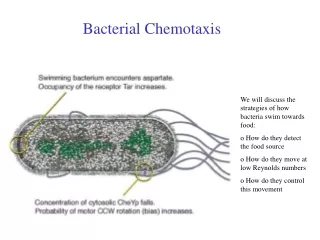
Near-Perfect Adaptation in Bacterial Chemotaxis. Yang Yang Advisor: Sima Setayeshgar Department of Physics Indiana University, Bloomington, IN. E. coli and Bacteria Chemotaxis. http://www.rowland.harvard.edu/labs/bacteria/index_movies.html.

E N D
Near-Perfect Adaptation in Bacterial Chemotaxis Yang Yang Advisor: Sima Setayeshgar Department of Physics Indiana University, Bloomington, IN Yang Yang, Candidacy Seminor
E. coli and Bacteria Chemotaxis http://www.rowland.harvard.edu/labs/bacteria/index_movies.html Increasing attractants or Decreasingrepellents Yang Yang, Candidacy Seminor
Chemotaxis Signal Transduction Network in E. coli Stimulus Signal Transduction Pathway [CheY-P] Motor Response Flagellar Bundling Histidine kinase Methylesterase Couples CheA to MCPs Response regulator Methyltransferase Dephosphorylates CheY-P Motion CheW CheR CheB CheZ CheA CheY Run Tumble Yang Yang, Candidacy Seminor
Robust Perfect Adaptation From Sourjik et al., PNAS (2002). Steady state [CheY-P] / running bias independent of value constant external stimulus (adaptation) Precision of adaptation insensitive to changes in network parameters (robustness) Adaptation Precison FRET signal [CheY-P] CheR fold expression Fast response Slow adaptation From Alon et al.,Nature (1999). Yang Yang, Candidacy Seminor
This Work: Outline • New computational scheme for determining conditions and numerical rangesfor parameters allowing robust (near-)perfect adaptation in the E. coli chemotaxis network • Comparison of results with previous works • Extension to other modified chemotaxis networks, with additional protein components • Conclusions and future work Yang Yang, Candidacy Seminor
Modified fine-tuned model • Ligand binding • Methylation • Phosphorylation phosphorylation methylation Ligand binding Yang Yang, Candidacy Seminor
Reaction rates Yang Yang, Candidacy Seminor
START with a fine-tuned model of chemotaxis network that: • reproduces key features of experiments • is NOT robust • AUGMENT the model explicitly with the requirements that: • steady state value of CheY-P • values of reaction rate constants, • are independent of the external stimulus, s, thereby explicitly incorporating perfect adaptation. : state variables : reaction kinetics : reaction rates : external stimulus Approach … Yang Yang, Candidacy Seminor
Augmented System The steady state concentration of proteins in the network satisfy: The steady state concentration of UN= [CheY-P] must be independent of stimulus, s: where parameter εallows for “near-perfect” adaptation. Reaction rates are constant and must also be independent of stimulus, s: Discretize s in range {slow, shigh} Yang Yang, Candidacy Seminor
Physical Interpretation of ε : Near-Perfect adaptation • Measurement of c = [CheY-P] by flagella motor constrained by diffusive noise • Relative accuracy*, • Signaling pathway required to adapt “nearly” perfectly, to within this lower bound • (*) Berg & Purcell, Biophys. J. (1977). : diffusion constant (~ 3 µM) : linear dimension of motor C-ring (~ 45 nm) : CheY-P concentration (at steady state ~ 3 µM) : measurement time (run duration ~ 1 second) Yang Yang, Candidacy Seminor
Implementation • Use Newton-Raphson (root finding algorithm with back-tracking), to solve for the steady state of augmented system, • Use Dsode (stiff ODE solver), to verify time- dependent behavior for different ranges of external stimulus by solving: Yang Yang, Candidacy Seminor
MichaelisMenten kinetics and constants A chemical reaction: A key assumption in this derivation is the quasi steady state approximation, namely that the concentration of the substrate-bound enzyme change much more slowly than those of the product and substrate and we can assume it is always in steady state, then: Where Km is the Michaelis Menten Constant(MM constant) Yang Yang, Candidacy Seminor
Converting from guess to solution Starting from initial guess A, the solver converted the solution to B A inverse of T3 M-M constant (K3R-1) B T3 autophosphorylation rate (k3a) Yang Yang, Candidacy Seminor
Parameter Surfaces Surface 2D projections Inverse of T1 methylation MM constant(k1R-1) Inverse of T1 demethylation MM constant(k1B-1) Inverse of T1 methylation MM constant(k1R-1) • 1%<<3% • 0%<<1% T1autophosphorylationrate K1a Yang Yang, Candidacy Seminork
Validation Verify steady state NR solutions dynamically using DSODE for different stimulus profiles: Concentration (µM) Time (s) Yang Yang, Candidacy Seminor
Violating and Restoring Perfect Adaptation 15% (5e+6,10) CheYp Concentration (µM) inverse of T3 M-M constant (K3R-1) 2% (1e+6,10) T3 autophosphorylation rate (k9) Time (s) Step stimulus from 0 to 1e-3M at t=500s Yang Yang, Candidacy Seminor
Results Conditions for (Near-)Perfect Adaptation Yang Yang, Candidacy Seminor
Inverse of Methylation MM constant Autophosphorylation Rate inverse of T1 M-M constant (K1R-1) inverse of T0 M-M constant (K0R-1) T0 autophosphorylation rate (k0a) T1 autophosphorylation rate (k1a) Yang Yang, Candidacy Seminor
Inverse of Methylation MM constant Autophosphorylation Rate(cont’d) inverse of T3 MM constant (K3R-1) inverse of T2 MM constant (K2R-1) T2 autophosphorylation rate (k2a) T3 autophosphorylation rate (k3a) Yang Yang, Candidacy Seminor
Inverse of Methylation MM constant Autophosphorylation Rate(cont’d) inverse of LT1 MM constant (K1LR-1) inverse of LT0 MM constant (K0LR-1) LT1 autophosphorylation rate (k1al) LT0 autophosphorylation rate (k0al) Yang Yang, Candidacy Seminor
Inverse of Methylation MM constant Autophosphorylation Rate(cont’d) inverse of LT3 MM constant (K3LR-1) inverse of LT2 MM constant (K2LR-1) LT3 autophosphorylation rate (k3al) LT2 autophosphorylation rate (k2al) Yang Yang, Candidacy Seminor
Inverse of Demethylation MM constant Autophosphorylation Rate inverse of T2 M-M constant (K2B-1) inverse of T1 M-M constant (K1B-1) T1 autophosphorylation rate (k1a) T2 autophosphorylation rate (k2a) Yang Yang, Candidacy Seminor
Inverse of Demethylation MM constant Autophosphorylation Rate(cont’d) inverse of T4 M-M constant (K4B-1) inverse of T3 M-M constant (K3B-1) T3 autophosphorylation rate (k3a) T4 autophosphorylation rate (k4a) Yang Yang, Candidacy Seminor
Inverse of Demethylation MM constant Autophosphorylation Rate(cont’d) inverse of LT2 MM constant (K2LB-1) inverse of LT1 MM constant (K1LB-1) LT2 autophosphorylation rate (k2al) LT1 autophosphorylation rate (k1al) Yang Yang, Candidacy Seminor
Inverse of Demethylation MM constant Autophosphorylation Rate(cont’d) inverse of LT4 MM constant (K3LB-1) inverse of LT3 MM constant (K2LB-1) LT4 autophosphorylation rate (k13) LT3 autophosphorylation rate (k12) Yang Yang, Candidacy Seminor
Methylation catalytic rate/demethylationcatlytic rate is constant T1methylationcatalytic rate T2methylationcatalytic rate T1demethylation catalytic rate T2demethylation catalytic rate Yang Yang, Candidacy Seminor
Methylation catalytic rate/demethylationcatlytic rate is constant T2methylationcatalytic rate T3methylationcatalytic rate T3demethylation catalytic rate T4demethylation catalytic rate Yang Yang, Candidacy Seminor
Methylation catalytic rate/demethylationcatlytic rate is constant LT0methylationcatalytic rate LT1methylationcatalytic rate LT1demethylation catalytic rate LT2demethylation catalytic rate Yang Yang, Candidacy Seminor
Methylation catalytic rate/demethylationcatlytic rate is constant LT2demethylation catalytic rate LT3demethylation catalytic rate LT3demethylation catalytic rate LT4demethylation catalytic rate Yang Yang, Candidacy Seminor
Summary • The Inverse of Methylation MM constants linearly decrease with Autophosphorylation Rates • The Inverse of Demethylation MM constants linearly increase with Autophosphorylation Rates • The ratio of Methylation catalytic rates and demethylation catlytic rates for the next methylation level is constant for all methylation states These conditions are consistent with those obtained in previous works from analysis of a detailed, two-state receptor model*. * B. Mello et al. Biophysical Journal , (2003). Yang Yang, Candidacy Seminor
Conditions in two-state receptor model • Receptor autophosphorylation rates are proportional to the receptor activity: • Only the inactive or active receptors can be methylated or demethylated. The association rates between receptors and CheR or CheBp are linearly related to the receptor activity, while dissociation rates are independent with 𝜆. Then the inverse of the methylation or demethylation MM constants are linearly related to the receptor activity: • The ratios between methylation catalytic rates and demethylation catalytic rates for the next methylation level are constant: • The phosphate transfer rates from CheA to CheB or CheY are proportional to receptor activities: Yang Yang, Candidacy Seminor
Results Conditions of protein concentrations for (Near-) Perfect Adaptation Yang Yang, Candidacy Seminor
Protein concentrations Yang Yang, Candidacy Seminor
Relationship between protein concentrations (M) (M) (M) (M) Yang Yang, Candidacy Seminor
Relationship between protein concentrations (M) (M) (M) (M) Yang Yang, Candidacy Seminor
Relationship between protein concentrations (M) (M) (M) (M) Yang Yang, Candidacy Seminor
Diversity of Chemotaxis Systems In different bacteria, additional protein components as well as multiple copies of certain chemotaxis proteins are present. Response regulator Phosphate “sink” Eg.,Rhodobacter sphaeroides, Caulobacter crescentus and several rhizobacteriapossess multiple CheYs while lacking of CheZ homologue. CheY2 CheY1 Yang Yang, Candidacy Seminor
Two CheY System Exact adaptation in modified chemotaxis network with CheY1, CheY2 and no CheZ: CheY1p (µM) CheY1p (µM) • Requiring: • Faster phosphorylation/autodephosphorylation rates of CheY2 than CheY1 • Faster phosphorylation rate of CheB Time(s) Yang Yang, Candidacy Seminor
Conclusions • Successful implementation of a novel method for elucidating regions in parameter space allowing precise adaptation • Numerical results for (near-) perfect adaptation manifolds in parameter space for the E. coli chemotaxis network, allowing determination of • conditions required for perfect adaptation, consistent with and extending previous works • numerical ranges for unknown or partially known kinetic parameters • Extension to modified chemotaxis networks, for example with no CheZ homologue and multiple CheYs Yang Yang, Candidacy Seminor
Future Work • Extension to other signaling networks • vertebrate phototransduction • mammalian circadian clock • allowing determination of • parameter dependences underlying robustness • plausible numerical values for unknown network parameters Yang Yang, Candidacy Seminor
vertebrate phototransduction • cGMP: cyclic GMP • PDE: cGMP phosphodiesterase • GCAP: guanylyl cyclase activating, Ca2+ binding protein • gc: guanylyl cyclase, which synthesis cGMP http://www.fz-juelich.de/inb/inb-1/Photoreception/ Yang Yang, Candidacy Seminor
Light adaptation of primate cones J. M. Valkkin and Dirk Van Norren,Vision Res. (1983). Yang Yang, Candidacy Seminor
Differential equations for verterbratephototransduction Russell D. Hamer, Visual Neuroscience (2000) Yang Yang, Candidacy Seminor
Mammalian circadian clock http://www.umassmed.edu/neuroscience/faculty/reppert.cfm?start=0 Yang Yang, Candidacy Seminor