Download
1 / 31
380 likes | 793 Views
Chap. 3A Amino Acids, Peptides, and Proteins. Topics Amino acids Peptides and proteins Working with proteins The structure of proteins: primary structure. Fig. 3-6. Absorption of ultraviolet light by aromatic amino acids. Introduction to Proteins.

E N D
Chap. 3A Amino Acids, Peptides, and Proteins • Topics • Amino acids • Peptides and proteins • Working with proteins • The structure of proteins: • primary structure Fig. 3-6. Absorption of ultraviolet light by aromatic amino acids.
Introduction to Proteins Proteins mediate nearly every process that takes place inside a cell. They are the most abundant biological macromolecules in cells. All proteins, regardless of organism, are composed of the same set of 20 amino acids that are incorporated into them during translation. Due to the nearly limitless variety in the sequences of amino acids in proteins, nearly all imaginable functions can be encoded in proteins (Fig. 3-1).
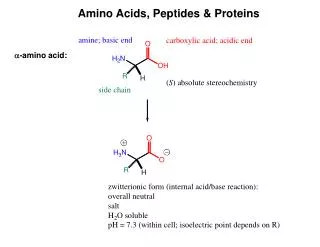
Common Features of Amino Acids There are 20 "standard" amino acids that are specified by the genetic code and polymerized into proteins by ribosomal translation. All amino acids contain an a carbon to which typically 4 different substituent groups are attached (Fig. 3-2). These groups are the a-amino group, the a-carboxyl group, hydrogen, and the variable R group (side-chain). (Note that in glycine, the R group consists of another hydrogen atom. In the other 19 amino acids the carbon is a chiral center.) The a-amino and a-carboxyl groups are charged at neutral pH. There are two possible configurations for these four substituents--the "D" and "L" stereoisomers, which are mirror images of each other (enantiomers) (Fig. 3-3). The standard amino acids have the L-configuration. Amino acids are classified based on the characteristics of their R groups. The chemical properties of the standard 20 amino acids are summarized in Table 3-1 (next two slides).
Numbering of Carbons in Amino Acids The conventions for labeling the carbon atoms in amino acids is illustrated using lysine in the figure. The carbon is always carbon-2 of the amino acid. The -carboxyl group is always carbon-1
D,L Classification System for Amino Acid Configurations The D, L system is used to specify the absolute configuration of substituent groups about chiral carbons in amino acids and monosaccharides. In this system, amino acids are specified as D or L based on comparison of their configurations to the reference compounds, D- and L-glyceraldehyde (Fig. 3-4). Note that not all amino acids actually are levorotatory. Thus D and L designate only the configurations of the substituent groups and not the optical properties of the amino acid.
Classification of Amino Acids by R Group Amino acids are collected into different categories based on similarities in the properties of their R groups. One such classification scheme (Fig. 3-5) relies heavily on the polarity of the R groups. Note that some amino acids such as glycine, histidine, and cysteine do not fit perfectly into any one group. Thus, their assignments to a particular group are somewhat subjective rather than based on absolute characteristics. The textbook places the amino acids into five categories based on the properties of their R groups--nonpolar, aliphatic; aromatic; polar, uncharged; positively charged; and negatively charged (next slides).
Nonpolar, Aliphatic Amino Acids The amino acids in this group lack polar functional groups in their side chains. Due to the hydrophobicity of their R groups, they often cluster together within the interior of proteins, stabilizing protein structure via hydrophobic interactions. The preferences of several of these amino acids for regions of protein secondary structure are discussed in Chap. 4.
Aromatic Amino Acids The side-chains of the aromatic amino acids, phenylalanine, tyrosine, and tryptophan, overall are very hydrophobic. The R group of tyrosine also contains a polar hydroxyl group that can participate in H bonding interactions.
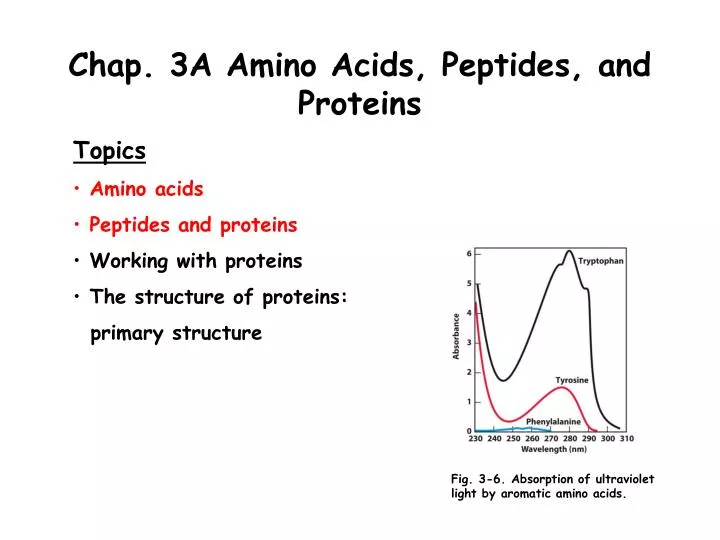
Aromatic Amino Acids and Protein Quantification The R groups of tyrosine, and particularly tryptophan, absorb ultraviolet light at a maximum of 280 nm wavelength (Fig. 3-6). Light absorption by these amino acids is exploited in detecting and quantifying proteins in the laboratory using the technique of absorbance spectrometry. The general configuration of a spectrophotometer is shown in the diagram in Box 3-1. The amount of a light-absorbing molecule in solution is determined from the Lambert-Beer law, A = cl. In this equation, the absorbance A (log I0/I) is equivalent to the product of the extinction coefficient (), times the solute concentration (c), times the path length of the solution (l).
Polar, Uncharged Amino Acids The R groups of the polar, uncharged amino acids all contain polar functional groups that can hydrogen bond with water. Serine and threonine contain hydroxyl groups, asparagine and glutamine contain amide groups, and cysteine contains a sulfhydryl group, albeit whose polarity is quite weak. Asparagine and glutamine are the amide forms of the two negatively charged amino acids aspartate and glutamate. The sulfhydryl group of the cysteine side chain is a weak acid (pKa = 8.2). The cysteine side chain therefore is mostly uncharged at neutral pH.
Cysteine and Disulfide Bonds The thiol groups of two cysteine residues are readily oxidized to form a covalently linked dimeric amino acid known as cystine. In cystine, the two cysteines are joined by a disulfide bond (Fig. 3-7). The disulfide-linked cystine residue is strongly hydrophobic. In proteins, disulfide bonds form covalent links between different parts of a polypeptide chain, or between two different polypeptide chains.
Positively Charged Amino Acids The most hydrophilic R groups are those that are either positively or negatively charged. The side-chains of lysine and arginine are fully positively charged at neutral pH. In lysine, a primary amino group is attached to the carbon of the side-chain. In arginine, the guanidinium group of the side-chain is postively charged. The histidine R group contains an aromatic imidazole group that is partially positively charged at neutral pH (Refer to Worked Example 2-5). Histidine residues function in many enzyme-catalyzed reactions as proton donors and/or acceptors.
Negatively Charged Amino Acids The R groups of aspartate and glutamate contain carboxyl groups that are fully negatively charged at neutral pH (pKRs of 3.65 and 4.25, respectively, Table 3-1). In aspartate, the carboxyl group is attached to the ß carbon of the amino acid backbone. In glutamate, the carboxyl group is attached to the carbon.
Uncommon Amino Acids (I) Many amino acids other than the standard 20 used in protein synthesis occur in nature. Some of these are derived by modification of amino acids already incorporated in a polypeptide (e.g., 4-hydroxyproline of collagen, and -carboxyglutamate of prothrombin, Fig. 3-8a).
Uncommon Amino Acids (II) Other amino acids in proteins are transiently modified as part of the control of the activity of the protein. Phosphorylation of the amino acids serine, threonine, and tyrosine is a particularly common modification of this kind (Fig. 3-8b).
Uncommon Amino Acids (III) Some 300 additional amino acids have been found in cells, many of which are not constituents of proteins. The amino acids ornithine and citrulline, for example, are intermediates in the biosynthesis of arginine and also are metabolites of the urea cycle (Fig. 3-8c)
Intro. to Amino Acid Ionization (I) The -carboxyl and -amino groups of all amino acids, along with the ionizable R groups of 7 amino acids, function as weak acids and bases in aqueous solutions (Table 3-1). The pKas of these functional groups depend on the chemical properties of the groups themselves and range between 1.8-2.4 for the -carboxyl groups, and 8.8-11.0 for -amino groups. When a simple amino acid such as alanine, which lacks an ionizable functional group in its side-chain, is dissolved in water at neutral pH, its -carboxyl group is negatively charged, and its -amino group is positively charged. Such dipolar ions (total charge equals 0) are called zwitterions. Substances having this dual (acid-base) nature are amphoteric and are often called ampholytes. The ionization behavior of the -carboxyl and -amino groups of a simple amino acid such as alanine are shown in the diagram below.
Intro. to Amino Acid Ionization (II) The nonionic and zwitterionic forms of a simple amino acid such as alanine are shown in Fig. 3-9. The zwitterionic form predominates at neutral pH. The nonionic form does not occur in significant amounts in aqueous solution at any pH. A zwitterion can act as either an acid (proton donor) or a base (proton acceptor).
Titration of Simple Amino Acids The titration curves of simple amino acids such as glycine (Fig. 3-10) that have non-dissociable R groups, have two plateaus, which correspond to the dissociation and titration of the -carboxyl group (pK1, left) and the -amino group (pK2, right). As shown above the curve, the predominant ionic species in solution at low pH is the fully protonated form, +H3N-CH2-COOH (net charge = +1), In between the two plateaus, the zwitterionic form, +H3N-CH2-COO- (net charge = 0) predominates. At the end of the titration, the fully dissociated species H2N-CH2-COO- (net charge -1) predominates. The curve shows that glycine has two regions of buffering power centered ±1 pH unit above pK1 and pK2. The Henderson-Hasselbalch equation can be used to calculate the amounts of the conjugate acid and conjugate base species in solution at any pH. Lastly, the pH at which the zwitterionic (0-charged) species of glycine predominates (one equivalent of OH- added) is called the isoelectric point or isoelectric pH. The isoelectric pH is exactly halfway between the two pKas for glycine.
Chemical Environment and the pKa The pKas of the -carboxyl groups of all amino acids are lower than the pKas of the carboxyl groups in methyl-substituted carboxylic acids such as acetic acid (Fig. 3-11). This is due to the local chemical environment of the -carboxyl groups in amino acids. Namely, placement near the -amino group, which is positively charged, makes the -carboxyl groups of amino acids more acidic than the carboxyl group of acetic acid. Similarly, the chemical environment near -amino groups makes them more acidic than the amino groups of a methyl-substituted amino compounds such as methylamine. In this case the electron withdrawing properties of the oxygens on the -carboxyl groups of amino acids make the -amino groups hold onto their protons less tightly than in other environments.
Titration of Glutamate The acidic amino acid, glutamate, has a second carboxyl group present in its side-chain. Thus the titration curve for glutamate (and aspartate) has three plateaus, each one corresponding to the dissociation of a proton from the amino acid (Fig. 3-12a). Since the R group carboxyl group has a pKR between that of the pK1 and pK2, the second plateau corresponds to the titration of this group. Based on inspection of the ionic forms in solution (top) it is clear that the zwitterionic form of glutamate occurs at a pH midway between that of pK1 and pKR. Thus the pI for glutamate is 3.22.
Titration of Histidine Histidine has an imidazole R group that contains a dissociable proton with a pKR of 6.0. Thus the titration curve for histidine also has three plateaus (Fig. 3-12b). For histidine, the 0-charged zwitterionic species occurs in solution at a pH midway between pKR and pK2 (pI = 7.59). Because the histidine pKR is near neutrality, the R group of histidine plays a role in buffering the pH of solutions containing proteins.
Peptide Bonds Peptide bonds are amide linkages that join amino acids in oligopeptides, polypeptides, and proteins. Peptide bonds are formed by condensation reactions in which the elements of water are removed (dehydration) from the reacting -amino and -carboxyl groups that come together to form the bond (Fig. 3-13). Note that peptide bonds in proteins are formed in vivo via a different mechanism than the one shown in the diagram. The term oligopeptide refers to polymers with relatively few amino acid residues. The term polypeptide signifies polymers of generally less than 10,000 mw, whereas the term proteins refer to longer polymers.
Structure of (Oligo)peptides The structure of the pentapeptide, serylglycyltyrosylalanylleucine (Ser-Gly-Tyr-Ala-Leu, SGYAL) is shown in Fig. 3-14. Note that all five amino acids are linked by peptide bonds (shaded groups). R groups are shown in red. When an amino acid sequence of a peptide, polypeptide, or protein is shown, by convention the amino-terminal (N-terminal) end is placed on the left, and the carboxyl-terminal (C-terminal) end is place on the right. The amino acid sequence is read left-to-right beginning with the N-terminal end.
Ionization Behavior of Peptides The -amino and -carboxyl groups of amino acid residues that are joined together in peptide bonds do not undergo ionization reactions near physiological pH (Fig. 3-15). The free -amino group at the N-terminus of the peptide, and the free -carboxyl group at the C-terminus ionize as they do in isolated amino acids. Likewise the R groups that contain ionizable functional groups continue to ionize as in free amino acids. Note, however, that the pKa values for ionizable residues in peptides change somewhat from their values in free amino acids due to changes in their local chemical environments. Like free amino acids, peptides have characteristic titration curves and an isoelectric point at which the net charge on the peptide is zero. These properties are exploited in some of the separation techniques discussed later in the chapter.
Aspartame Naturally occurring peptides range in length from two to many thousands of amino acid residues. Even small peptides can be biologically active. For example, the commercially synthesized dipeptide L-aspartyl-L-phenylalanine methyl ester is the sweetener called aspartame or NutraSweet. Other examples of short biologically active peptides are thyrotropin-releasing factor (three amino acid residues) and oxytocin (nine amino acid residues).
Protein Sizes The lengths of polypeptide chains in proteins vary considerably (Table 3-2). While the great majority of proteins contain fewer than 2,000 amino acids, some are much larger. The largest known protein is titin (26,926 amino acids), which is a component of vertebrate muscle. Some proteins consist of a single polypeptide chain, whereas others called multisubunit proteins have two or more associated polypeptides. The individual polypeptide chains in a multisubunit proteins can be identical or different. If at least two subunits are identical, the protein is said to be oligomeric and the identical units (consisting of one or more polypeptide chains) are referred to as protomers.
Amino Acid Compositions of Proteins The amino acid compositions of proteins differ between proteins and are highly variable (Table 3-3). Some amino acids occur only once or not at all in a given protein and other amino acids may occur in large numbers. Based on the frequencies at which the 20 standard amino acids occur in proteins, and their molecular weights (Table 3-1), the average molecular weight of an amino acid in a protein is 128. Because a molecule of water (Mr 18) is lost on creation of each peptide bond, the average molecular weight of an amino acid residue in a protein is about 110.
Conjugated Proteins Simple proteins, such as chymotrypsin, contain only amino acids. Other proteins--conjugated proteins--contain other associated chemical components in addition to amino acids. The non-amino acid part of a conjugated protein is called its prosthetic group. Examples of conjugated proteins and their prosthetic groups are listed in Table 3-4.