Download

1 / 37
370 likes | 498 Views
Prions as proteinaceous genetic material Michael D. Ter-Avanesyan. Michael D. Ter-Avanesyan. Yeast Sup35, transmission electron microscopy. β -2 microglobulin , Atomic force microscopy. Amyloids. Fibrils form via autocatalytic non-covalent protein polymerization, accompanied

E N D
Prions as proteinaceous genetic material Michael D. Ter-Avanesyan Michael D. Ter-Avanesyan
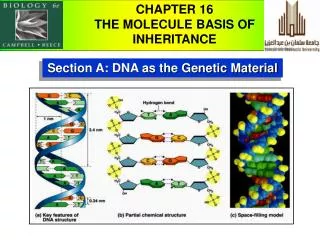
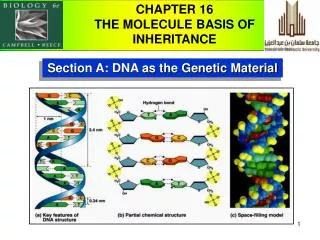
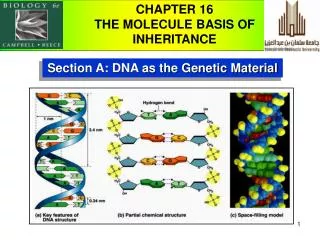
Yeast Sup35, transmission electron microscopy β-2 microglobulin, Atomic force microscopy Amyloids • Fibrils form via autocatalytic non-covalent protein polymerization, accompanied • by deep conformational rearrangement of polymerizing protein monomers • Have a specific cross-β structure, in which the β strands are perpendicular to • the fibril axis, while β sheets formed by separate monomers are parallel to it • Rigid, insoluble in detergents, resistant to proteases, bind Congo red and thioflavine T 2 Kajava et al., PNAS USA, 2004 Shewmakeret al., PNAS USA, 2006 2
Amyloid diseases Transmissible (prion) amyloidoses: Creutzfeldt–Jakob disease Gerstmann– Straussler–Scheinker disease Fatal familial insomnia Kuru Sheep scrapie Bovine spongiform encephalopathy Non-transmissible (non-prion) amyloidoses: Alzheimer disease Parkinson disease Huntington disease… ________________ ~40diseases Amyloid form of various unrelated proteins Infectious amyloids of the PrP protein 3
Emergence of mechanisms preventing protein aggregation and amyloidogenesis Primordial peptides prone to form amyloids (scaffold structures) Polypeptides evolved to acquire biologically relevant globular structure Cells developed aggregation-clearing mechanisms __________________________________________ Nowadays -amyloidogenic proteins can be both detrimental and beneficial 5
Amyloid-like polymers of ACS adaptor act as a platform for caspase activation LRR NLRP3 NBD NLPR3 PYD PYD ACS CARD ACS CARD Caspase-1 Caspase domain Caspase-1 filaments Lu et al., Cell, 2014 6
Role of amyloid in melanin polymerization (Pmel17 amyloid provides a scaffold for melanin synthesis) From: Inge-Vechtomov et al., Prion, 2007 7
In mammals prions are proteinaceous infectious agents Bolton D., McKinley M., Prusiner S. 1982. Identification of protein that purifies with the scrapie prion. Science, 218, 1309-1311 In lower eukaryotes prions are proteinaceous genetic material Wickner R, 1994. [URE3] as an altered URE2 protein: evidence for a prion analog in S. cerevisiae. Science, 264, 566-569 8
Central dogma of molecular biology (Information flow in biological systems) DNA Addition ____________ RNA Protein Protein Function Function F. Crick, Nature, 1970
[PSI+] determinant Cox, Heredity, 1965 (Mendel, 1865) • [PSI+] manifests a nonsense suppressor phenotype • [PSI+] is inherited in a non-Mendelian fashion and can be • transmitted from one cell to another with cytoplasm • No extrachromosomal DNA or RNA have been found to be • associated with [PSI+] • [PSI+] can be efficiently eliminated by protein denaturing agent, • guanidine hydrochloride, or by exposure to stress-inducing • factors • The curing of [PSI+] is reversible Hypothesis The [PSI+] phenotype reflects conversion of the Sup35 protein into a prion form related to its aggregation and functional inactivation Wickner, Science, 1994 11
* * * * The system for [PSI+] detection (Suppression of ade2 nonsense mutations) mRNA Wild type Translation mRNA Nonsense mutant Translation mRNA Suppression Translation 12
1 124 254 685 N M C mRNA ААА protein Sup45/35 Domain structure of the Sup35 protein Sup45 (eRF1)andSup35 (eRF3) –translation termination factors Rich in: Gln, Asn Glu, Lys Nonconservative Nonessential for translation termination and viability Structurally similar to translation elongation factor eEF1A Essential for viability OnlyNdomain is required for [PSI+] Ter-Avanesyan et al., Mol. Microbiol.,1993; Genetics,1994 13
Structural organization of the Sup35 fibril Paushkin et al., Mol. Cell. Biol.,1997 Baxa et al., Mol.Microbiol., 2011 14
[PSI+] pellet pellet pellet psi- psi- psi- psi- Sup35 is 400-fold mutiplicated in 4 cycles Prion inheritance in vitro Sup35 forms prion aggregates in [PSI+] cells Serial propagation of the [PSI+] prion Centrifugation soluble pellet ribosomes 1 2 3 4 5 6 7 8 9 10 11 12 [PSI+] Sup35 [psi-] Sup35[PSI+] is aggregated Paushkin et al., EMBO J., 1996; Science, 1997 15
Prion transformation of yeast (Proof of the prion concept) Sup35 polymers are infectious and underlie the [PSI+] determinant Tanaka, Weissman et al., Nature, 2004 (Avery et al., J. Exptl. Med., 1944) Sup35NM was produced in E. coli, purified and polymerizedin vitro Yeast spheroplasts was co-transformed with Sup35 fibrils and a plasmid carrying the URA3 selectable marker Transformation resulted in the appearance of clones with the [PSI+] phenotype (white color) 16
Role of chaperones in [PSI+] maintenance SSA1+ YDJ1 Control SSA1 YDJ1 SSB1 HSP104 [PSI+PS] Overproduction of Ssa1, Ssb1 (Hsp70/DnaK) and Ydj1(Hsp40/DnaJ) chaperones can destabilize [PSI+] [PSI+] Kryndushkin et al., J. Biol. Chem., 2002 17
Hsp104 plays a key role in [PSI+] maintenance Hsp104 [PSI+] cannot propagate in the absence of Hsp104 Paradox: Overproduction ofHsp104 also can cause [PSI+] loss Chernoff et. al., Science, 1995 To explain the role of Hsp104,we relied on two considerations: Sup35fibrils formed in vitro • Sup35 prion particles have fibrillar shape • Hsp104act on fibrils in the same way as on • aggregates of thermally denatured proteins, • i.e. disrupt them 18
Sup35: Hsp104 The model of prion replication Polymerization (only Sup35 is required) Fragmentation (Hsp104 is required) Hsp104-mediated fragmentation of prion particles multiplicates them which is necessary for their stable inheritance Kushnirov and Ter-Avanesyan, Cell, 1998 Paradox: Hsp104, which has evolved for destruction of protein aggregates is essential for maintenance and inheritance of prion aggregates 19
Methods of prion particles analysis • Isolation of aggregates by centrifugation • Microscopic observation of aggregated GFP hybrid proteins • Both methods do not allow to analyze the size of polymers GFP GFP [PSI+] [psi-] Sup35NM Patino, Lindquist, Science, 1996 20
Start kDa 4200 (titin) Stability of Sup35polymers in the presence of SDS 740 (nebulin) SDS, % 2 2 2 5 2 2 2 2 205 (myosin) Start Polymers Sup35 0 25 37 42 50 70 100 37 Sup35monomers Monomers Sup35 [psi-] [PSI+] Electrophoretic analysis of prion polymers Polymer SDS disrupts aggregates to polymers Analysis of polymersin agarose gel with SDS Kryndushkin et al., J. Biol. Chem., 2003 21
Hsp104 fragments prion polymers Both deletion of HSP104 and incubation of cells on medium with GuHCl cause loss of[PSI+]; GuHClinhibitsHsp104 activity Start Start kDa kDa 4200 4200 740 740 205 Sup35 monomer 205 210 100 55 30 17 11 8 % Hsp104 Sup35 мономер 0 1 2 3 4 5 6 Cell generations 0 1 2 3 1 2 3 Generations on GuHCl Hours after GuHCl Decrease in the level of Hsp104causes increase of the polymer size The size of Sup35 polymer is a characteristic trait of the [PSI+] prion Kryndushkin et al., J. Biol. Chem., 2003 22
[PSI+] variants (“alleles”) differ in suppressor phenotype and the size of Sup35 polymers 1 2 3 4 5 6 7 8 weak [PSI+] weak [PSI+] [psi -] weak [PSI+] prion variant: W S S S S W S S [PSI+] [PSIPS+] W - weak; S - strong strong [PSI+] strong [PSI+] Hypothesis: Sup35 polymers corresponding to different [PSI+] variants differ by susceptibility to fragmentation by Hsp104 23
Insertion of tyrosine residues into the polyglutamine stretch enhances efficiency of polymer fragmentation KDa (125) (254) (685) polyQ/QY M C 4000 Sup35MC polyQ: MSG-(QQQQQ)m-QSQGA polyQY: MSG-(QQQYQ)m-QSQGA polyQ/QY proteins 730 25Q 45Q 51Q 56Q 65Q 70Q 131Q 30QY 46QY 50QY 76QY 120QY Tyrosine residues stimulate fragmentation Alexandrov et al., J. Biol. Chem., 2008 Polymers inSDS-agarose gel 24
Non-Q/N residues in yeast prion domains Fragmentation efficiency Inhibit polymerization 25
Role of the exposed region of the Sup35 prion domain in fragmentation of polymers Hypothesis: Difference in [PSI+] phenotypes is related to variation in exposure of certain non-Q/N amino acids in prion domains Alexandrov et al., PLoS One, 2012 26
Role of chaperones in fragmentation of Sup35 prion polymers 27
Origin of prion proteins in yeast: hypothesis • Background: • PolyQ domains can expand and contract • PolyQ often serve to mediate interaction between proteins • Expansion of polyQ stretches can result in toxicity of corresponding proteins • while their contraction may inhibit biological function of these proteins • Insertionof non-Q/N amino acid residues into polyQ should stabilize their length • and may decrease toxicity • Hypothesis: • Yeast prion domains may have derived from polyQ tracts via accumulation • and amplification of mutations Alexandrov, Ter-Avanesyan ,Prion, 2013 28
Main contributors: V. Kushnirov S. Paushkin (now in USA) A. Alexandrov D. Kryndushkin (now in USA) I. Alexandrov (now in USA) Thank you for your attention 29
Soluble fraction Soluble fraction Pellet Pellet Sup35PS [PSI+PS] Sup35 [PSI+] Hsp104: Wild type level Enhanced level Excess of Hsp104 dissolves prion aggregates of Sup35 and decreases their size Kushnirov et al., EMBO J., 2000 17
Sup35 forms prion aggregates in [PSI+] cells Proteinase K 8.0 4.0 2.0 1.0 0.4 0.2 (mkg/ml) Centrifugation [PSI+] Sup35 soluble pellet [psi-] Sup35 ribosomes 1 2 3 4 5 6 7 8 9 10 11 12 [PSI+] [psi-] Sup35[PSI+] is persistent to proteinase К Sup35[PSI+] is aggregated Paushkin et al., EMBO J., 1996 13
[PSI+] pellet pellet pellet psi- psi- psi- psi- Sup35 is 400-fold mutiplicated in 4 cycles Prion inheritance in vitro Serial propagation of the [PSI+] prion Paushkin et al., Science, 1997 14
Origin of prion proteins in yeast: hypothesis • Background: • PolyQ domains can expand and contract • PolyQ often serve to mediate interaction between proteins • Expansion of polyQ stretches can result in toxicity of corresponding proteins • while their contraction may inhibit biological function of these proteins • Insertionof non-Q/N amino acid residues into polyQ should stabilize their length • and may decrease toxicity • Hypothesis: • Yeast prion domains could derive and evolve due to mutational insertion some • amino acid residues into polyQ and their subsequent amplification Alexandrov, Ter-Avanesyan ,Prion, 2013 38
Prions in lower eukaryotes can be considered as: • Molecular basis of inheritance of acquired traits • Proteinaceous genes • An addition to the central dogma of molecular biology 5
Prions as proteinaceous genes • How do proteinaceous genes determine phenotypic traits? • What kind of information they encode? • How do proteinaceous genes replicate? • What is the nature of their “alleles”? • What is the biological significance of prions • (are they a bate or benefit)? 7
[PSI+] variants (“alleles”) differ in suppressor phenotype and the size of Sup35 polymers weak [PSI+] weak [PSI+] [psi -] weak [PSI+] strong [PSI+] strong [PSI+] Hypothesis: Sup35 polymers corresponding to different [PSI+] variants differ by susceptibility to fragmentation by Hsp104 22