Download

1 / 97
980 likes | 1.37k Views
NBCR Summer Institute 2006: Multi-Scale Cardiac Modeling with Continuity 6.3 Tuesday: Modeling Cardiac Myocyte Excitation-Contraction Coupling and its Regulation Andrew McCulloch, Anushka Michailova and Stuart Campbell. Systems Physiology Models. Circulatory system dynamics.

E N D
NBCR Summer Institute 2006:Multi-Scale Cardiac Modeling with Continuity 6.3Tuesday:Modeling Cardiac Myocyte Excitation-Contraction Coupling and its RegulationAndrew McCulloch, Anushka Michailova and Stuart Campbell
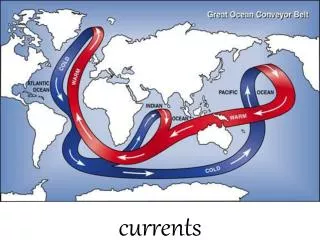
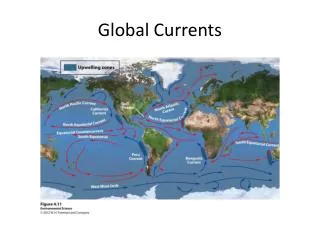
Systems Physiology Models Circulatory system dynamics Neurohumoral regulation Circulation Regulation of peripheral resistances, fluid volumes, HR Autonomic mediators Ventricular systolic pressures and cardiac output Coronary ostia pressure Torso bioelectric fields Torso Finite element and boundary element models Ventricular filling pressures and output impedance Epicardial potential fields Whole ventricular electromechanics Coronary artery flow Ventricles Transmembrane potentials Regional wall stresses, strains, displacements Coronary artery flows Intramyocardial pressure and volume Constitutive Models Tissue perfusion Wall stress and strain Action potential propagation Myocardium Total transmembrane ionic current Myofilament tension Crossbridge interactions Myofilament activation Ionic currents Calcium handling Cell Systems models Myocyte Cell signaling Purine metabolism adenosine Mitochondrial metabolism substrates
Multi-scale Modeling crossbridge lattice ventricles multicellular circulation
Tuesday: Modeling Myocyte Excitation-Contraction Coupling and its Regulation Cardiac mechanics: myofilament models Cardiac electrophysiology: ionic models Regulation of excitation-contraction coupling: biochemical models Intracellular Diffusion of Second Messengers – cAMP, Ca Getting set up with Continuity 6.3 A simple reaction diffusion model with Continuity 6.3
Wednesday: Finite Element Discretization and Mesh Fitting Finite element approximation and discretization Finite element interpolation – Lagrange and Hermite basis functions with Continuity 6.3 Tensor-product interpolation for two and three dimensions Curvilinear world coordinates and coordinate conversion Fibers and fields in Continuity 6.3 Least squares fitting of anatomic meshes and fiber angles with Continuity 6.3 Non-homogeneous strain analysis
Thursday: Monodomain Modeling in Cardiac Electrophysiology Cardiac myocyte ionic models Modeling cardiac action potential propagation Collocation FEM for monodomain problems with Continuity 6.3
Friday: Cardiac Mechanics and Electromechanics • Modeling Ventricular Wall Mechanics • Determinants of ventricular wall stress • Galerkin FEM for ventricular stress analysis • Systolic wall strains and anisotropy • Ventricular Electromechanics • Cardiac resynchronization therapy • Ventricular-Vascular Coupling
Cardiac Myocytes • Rod-shaped • Striated • 80-100 m long • 15-25 m diameter
Myocyte Connections • Myocytes connect to an average of 11 other cells (half end-to-end and half side-to-side) • Functional syncytium • Myocytes branch (about 12-15º) • Intercalated disks • gap junctions • connexons • connexins
endocardium midwall epicardium Fiber-Sheet Structure x510
Myocyte Ultrastructure • Sarcolemma • Mitochondria (M) ~30% • Nucleus (N) • Myofibrils (MF) • Sarcoplasmic Reticulum and T-tubule network
Striated Muscle Ultrastructure Electron micrograph of longitudinal section of freeze-substituted, relaxed rabbit psoas muscle. Sarcomere shows A band, I band, H band, M line, and Z line. Scale bar, 100 nm. From Millman BM, Physiol. Rev. 78: 359-391, 1998
The Sarcomere AnisotropicIsotropic to polarized light ~ 2.0 m
Excitation-Contraction Coupling • Calcium-induced calcium release • Calcium current • Na+/Ca2+ exchange • Sarcolemmal Ca2+ pump • SR Ca2+ ATP-dependent pump Click image to view animation of calcium cycling http://www.meddean.luc.edu/lumen/DeptWebs/physio/bers.html
Isometric Tension in Skeletal Muscle:Sliding Filament Theory (a) Tension-length curves for frog sartorius muscle at 0ºC (b) Developed tension versus length for a single fiber of frog semitendinosus muscle
700 500 200 300 400 600 100 Isometric Testing Sarcomere length, mm Sarcomere isometric 2.1 2.0 Muscle isometric 1.9 Tension, mN 2.0 1.0 time, msec
Isometric Length-Tension Curve Peak developed isometric twitch tension (total-passive) High calcium Low calcium sarcomere isometric muscle isometric Passive
Length-Dependent Activation Isometric peak twitch tension in cardiac muscle continues to rise at sarcomere lengths >2 mm due to sarcomere-length dependent increase in myofilament calcium sensitivity
Isotonic Testing Isovelocity release experiment conducting during a twitch Cardiac muscle force-velocity relation corrected for viscous forces of passive cardiac muscle which reduce shortening velocity
Modeling Myofilament Force Production • Ca2+ binding to TnC causes tropomyosin to change to a permissive state • Force development occurs as actin-myosin crossbridges form • Crossbridges can ‘hold’ tropomyosin in the permissive state even after Ca2+ has dissociated
Ca2+ Ca2+ Myofilament Activation/Crossbridge Cycling Kinetics Ca2+ kn Roff Roff Non-permissive Tropomyosin 0 * kb Ca2+ * koff kon koff Permissive Tropomyosin Ron Ron 0 * g g f f Ca2+ Permissive Tropomyosin, 1-3 crossbridges attached (force generating states) 0 A1 A1 * Ca2+ Ca2+ not bound to TnC Ca2+ bound to TnC This scheme is used to find A(t), the time-course of attached crossbridges for a given input of [Ca2](t)
Myofilament Model Equations • Total force is the product of the total number of attached crossbridges, average crossbridge distortion, and crossbridge stiffness: • Average crossbridge distortion is obtained by the solution to the following differential equation:
Passive Biaxial Properties 1 0 Fiber stress 8 6 Stress (kPa) 4 Cross-fiber stress 2 0 0. 0 0 0. 0 5 0. 1 0 0. 1 5 0. 2 0 0. 2 5 Equibiaxial Strain
Strain Energy • W is the strain-energy function; its derivative with respect to the strain is the stress. • This is equivalent to saying that the stress in a hyperelastic material is independent of the path or history of deformation. • Similarly, when a force vector field is the gradient of a scalar energy function, the forces are said to be conservative; they work they do around a closed path is zero. • The strain energy in an elastic material is stored as internal energy or free energy (related to entropy) …
Cauchy’s formula:t(n)= n•T In index notation: T33 e3 n T32 S a T31 T23 R T13 T22 T12 T21 T11 e2 Tij= ti•ej e1 Cauchy Stress Tensor is Eulerian Tij is the component in the xj direction of the traction vector t(n) acting on the face normal to the xi axis in the deformed state of the body. The "true" stress.
Lagrangian Stress Tensors The (half) Lagrangian Nominalstress tensorS SRj is the component in the xj direction of the traction measured per unit reference area acting on the surface normal to the (undeformed) XR axis. Useful experimentally S = detF.F-1.T ST The symmetric (fully) Lagrangian Second Piola-Kirchhoff stress tensor • Useful mathematically but no direct physical interpretation • For small strains differences between T, P, S disappear
A L F F a l Example: Uniaxial Stress undeformed length = L undeformed area = A deformed length = l deformed area = a Cauchy Stress Nominal Stress Second Piola-Kirchhoff Stress
Hyperelastic Constitutive Law for Finite Deformations Second Piola-Kirchhoff Stress Cauchy Stress
2-D Example:Exponential Strain-Energy Function Stress components have interactions
3-D Orthotropic Exponential Strain-Energy Function From: Choung CJ, Fung YC. On residual stress in arteries. J Biomech Eng 1986;108:189-192
Strain Energy Functions Transversely Isotropic (Isotropic + Fiber) Exponential Transversely Isotropic Exponential Transversely Isotropic Polynomial Orthotropic Power Law
Incompressible Materials Stress is not completely determined by the strain because a hydrostatic pressure can be added to Tij without changing CRS. The extra condition is the kinematic incompressibility constraint To avoid derivative of W tending to p is a Lagrange multiplier (a negative stress)
Extracellular Ion Concentrations Intracellular Ion Concentrations Resting Membrane Potential • An imbalance of total ionic charge leads to a potential difference across a cell membrane: Resting Membrane Potential Vm = Vo - Vi • Only a slight imbalance is needed to result in a potential difference • If 1/100,000th of available cytosolic K+ ions crossed the membrane of a spherical cell 10µm DØ, the membrane potential changes by 100 mV
3 Time (msec) Potential (mV) 4 2 threshold voltage 1 5 Ventricular Action Potential • Through a stimulus such as current injection, Vm reaches a threshold voltage • Voltage-gated Na+ ion channels change to open state, Na+ ions enter depolarizing the cell • A peak is reached near the equilibrium potential for Na+ ions • Other ion channels react, namely K+ and Ca2+ channels repolarizing the cell • Over time the cell returns to a resting equilibrium state
Inactivated Open Closed Na+ 3Na2+ ATP 2K+ Ion Motion • Ions cross the membrane by two methods: Active Transport and Diffusion Nernst Equation Ion transporters, including ligand- and voltage-gated channels, exchangers and ATP-dependent pumps V Potential Co Concentration of ion outside cell Ci Concentration of ion inside cell R Gas Constant z Valence F Faraday’s constant At 37°C, RT/F = 26 mV
Electrochemical Equilibrium • Ions in a resting cell are in electrochemical equilibrium Goldman-Hodgkin-Katz Equation Vm Membrane Potential Pion Permeability of membrane to particular ion [C]ion Concentration of a particular ion Requires the assumption of a constant electric field in the membrane
Voltage Clamping • Whole cell and patch clamping techniques reveal ionic current activity • Allows control over membrane voltage • And ionic concentrations on both sides of the membrane • Hodgkin and Huxley observed sodium and potassium currents in nerve cells • Hodgkin, A.L. and A.F. Huxley, A quantitative description of membrane current and its application to conduction and excitation in nerve. J Physiol, 1952. 117: p. 500-544.
Hodgkin-Huxley Ionic Currents Ohm’s Law: Nernst potentials Conductances = 1/resistivity where and n, m, and h are gating variables
n is the gating variable for the K+ current n = #open/(#open+#closed) = fraction of open channels = probability a channel is open αn = rate of channel opening βn = rate of channel closing αnand βn are f(Vm) found by fitting Potassium Gating (1) Potassium conductance was found empirically to have behavior: Corresponds to four protein subunits with equal open probability. Exponent represents # of gates per channel
Potassium Gating (2) (1) Solution of (1) neglecting αnand βn is: where no is an initial condition, and n∞ comes from steady state: So (1) can be written: