Download

1 / 28
280 likes | 404 Views
A Non-EST-Based Method for Exon-Skipping Prediction. Rotem Sorek, Ronen Shemesh, Yuval Cohen, Ortal Basechess, Gil Ast and Ron Shamir Genome Research August 2004 楊佳熒. Segments and blocks >300kb in size with conserved in human are superimposed on the mouse genome.

E N D
A Non-EST-Based Method for Exon-Skipping Prediction Rotem Sorek, Ronen Shemesh, Yuval Cohen, Ortal Basechess, Gil Ast and Ron Shamir Genome Research August 2004 楊佳熒
Segments and blocks >300kb in size with conserved in human are superimposed on the mouse genome Homologous human and mouse exon are, on the average, 85% identical in their sequences, but introns are more pooly conserved. (Waterston et al. Nature,2002)
Reference • Sorek, R.et al.Intronic Sequences Flanking Alternatively Spliced Exons Are Conserved Between Human and Mouse. Genome Research, 2003. • Sorek, R. et al. How prevalent is functional alternative splicing in the human genome. TRENDS in Genetics, 2004. • Sorek, R. et al.A Non-EST-Based Method for Exon-Skipping Prediction. Genome Research, 2004.
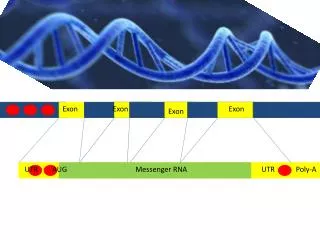
What is Exon-Skipping ? gene exon1 exon2 exon3 exon4 exon5 exon6 est1 est2 est3 est4 dbESTs
Intronic Sequences Flanking Alternatively Spliced Exons Are Conserved Between Human and Mouse Rotem Sorek and Gil Ast Genome Research July 2003
Objective and Result 1. Alternatively spliced conserved exons Human gene exon1 A1 exon2 B1 exon3 Alternatively spliced internal exons Human est1 3583 Human est2 Alternatively spliced conserved exons Mouse est1 243 Mouse est2 Mouse gene exon1 A2 exon2 B2 exon3 223/243=92% 199/243=82% 188/243=77% 2. Constitutively spliced conserved exons Human gene exon1 C1 exon2 D1 exon3 Constitutively spliced internal exons Human est1 7557 Human est2 Constitutively spliced conserved exons Human est3 1966 Human est4 Mouse est Mouse gene exon1 C2 exon2 D2 exon3 886/1966=45% 691/1966=35% 343/1966=17%
Per-position conservation near alternatively and constitutively spliced exons
KCDN3 gene exon information
KCDN3 gene exon 6 sequences (bold) (alternatively spliced exon)
Review : Finding exon-skipping events that are conserved between human and mouse 243 Conserved exon skipping events (25%) 737(980-243) Non-Conserved exon skipping events(75%)
How prevalent is functional alternative splicing in the human genome ? Rotem Sorek, Ron Shamir and Gil Ast TRENDS in Genetics Vo1.20 February 2004
Motivation • How many of there predicted splice variants are functional? • How many are the result of aberrant splicing (noise data)?
The influence of alternatively spliced exon on the protein-coding sequence. are peptide cassettess are peptide cassettess
Features differentiating between conserved alternatively spliced exons and non-conserved alternatively spliced exons 30% 62%
Conclusion • We show that conserved (functional) cassette exons possess unique characteristics in size, repeat content and in their influence on the protein. • By contrast, most non-conserved cassette exons do not share these characteristics. • We conclude that a portion of skipping exon evidence in EST databases is not functional, and might result from aberrant rather than regulated splicing.
Review :Intronic Sequences Flanking Alternatively Spliced Exons Are Conserved Between Human and Mouse 1. Alternatively spliced conserved exons Human gene exon1 A1 exon2 B1 exon3 Alternatively spliced internal exons Human est1 3583 Human est2 Alternatively spliced conserved exons Mouse est1 243 Mouse est2 Mouse gene exon1 A2 exon2 B2 exon3 223/243=92% 199/243=82% 188/243=77% 2. Constitutively spliced conserved exons Human gene exon1 C1 exon2 D1 exon3 Constitutively spliced internal exons Human est1 7557 Human est2 Constitutively spliced conserved exons Human est3 1966 Human est4 Mouse est Mouse gene exon1 C2 exon2 D2 exon3 886/1966=45% 691/1966=35% 343/1966=17%
Review : Features Differentiating Between Alternatively Spliced and Constitutively Spliced Exons
A Non-EST-Based Method for Exon-Skipping Prediction Rotem Sorek, Ronen Shemesh, Yuval Cohen, Ortal Basechess, Gil Ast and Ron Shamir Genome Research August 2004
Objective • Our goal was to find a combination of features that would detect a substantial fraction of the alternative exons. • The features we have chosen are the following : • exon length • divisible / not divisible by 3 • percent identity when aligned to the mouse • conservation in the upstream and downstream intronic sequences
Result • The best rule is : • at least 95% identity with mouse exon counterpart • exon size is a multiple of three • a best local alignment of at least 15 intronic nucleotides upstream of the exon with at least 85% identity • a perfect match of at least 12 intronic nucleotides downstream of the exon • The combination of features identified 76 exons, 31% of the 243 alternatively spliced exons in the training sets, whether non of 1753 constitutively spliced exons matched these features.
To test this classifier in a genome-wide manner (cont.) 108,983 human exons for which a mouse counterpart could be identified 108,983 using these rules 952 candidate exon, ~1%, were found. • For 453(48%) of the 952 candidate alternative exon there was such • skipping evidence. • Only(17%) of the 453 exons that were classified by our rule had their • exon-skipping supported by only one EST. • 3. The rest were supported by two or more.
To test this classifier in a genome-wide manner (cont.) 108,983 human exons for which a mouse counterpart could be identified 108,983 7% (7495 exons) out of our entire set search ESTs and cDNA • In comparison, skipping was supported by only a single EST in 46% of the • total 7495 exons. • This suggests that our classification rule enriches for alternatively spliced • exons with higher probability of being “real” relative to alternative exons • merely supported by EST evidence.
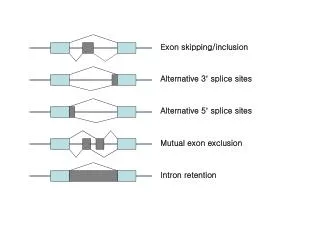
To test this classifier in a genome-wide manner • The remaining 499 candidate alternative exons (952-453) for which no EST/cDNA showing an exon skipping event was found. • Using the UCSC genome browser to check, we found that for 190 additional exons there was a human expressed sequence showing patterns of alternative splicing other than exon skipping cases. • Alternative donor/acceptor 22% • Intron retention17% • Mutually exclusive exon 7% • Thus, for 643(453+190 ; 68%) of the 952 candidate alternative exons identified by this method, there was independent evidence for alternative splicing in dbEST.
Conclusion • We show that a substantial fraction of the splice variants in the human genome could not be identified through current human EST or cDNA data. • In the future, we hope it could develop into a more general alternative splicing predictor that would identify other types of alternative splicing.
Classification of alternative splicing • Skipped Exons 2. Multiple Skipped Exons 3. Alternative Donor / Acceptors 4. Retained Introns