Download

1 / 50
510 likes | 836 Views
第 34 章、 DNA 复制. 34.1 DNA 复制是半保留式的 34.2 DNA 复制是双向进行的 34.3 DNA 聚合酶 III 催化复制叉处的聚合反应 34.4 DNA 聚合酶 III 同时催化两条链的合成 34.5 复制叉移动需要多蛋白复合物 34.6 DNA 复制起始于细菌染色体上的唯一的一个部位 34.7 DNA 复制终止于 ter 区 34.8 DNA 复制的其它方式 34.9 损伤的 DNA 可以修复. 遗传中心法则. DNA 复制. RNA 复制. 转录. 翻译. 蛋白质. 逆转录.

E N D
第34章、 DNA复制 34.1 DNA复制是半保留式的 34.2 DNA复制是双向进行的 34.3 DNA聚合酶III催化复制叉处的聚合反应 34.4 DNA聚合酶III同时催化两条链的合成 34.5 复制叉移动需要多蛋白复合物 34.6 DNA复制起始于细菌染色体上的唯一的一个部位 34.7 DNA复制终止于ter区 34.8 DNA复制的其它方式 34.9 损伤的DNA可以修复
遗传中心法则 DNA复制 RNA复制 转录 翻译 蛋白质 逆转录
34.1 DNA复制方式是半保留式 Meselson- Stahl实验: 1、使E.coli在含有唯一氮源15N(15NH4Cl)的培养基中培养,合成的所有核苷酸都含有15N ,具有较高的密度,都整合到亲代DNA中。 2、将生长在15NH4Cl培养基的E.coli转移到含有唯一氮源,但密度较低的14N(14NH4Cl)培养基中培养。培养两代,并分别取每一代的DNA进行密度梯度离心。
E.coli 细胞 首先在15NH4Cl培养基中培养 然后转到14NH4Cl培养基中培养 只在15NH4Cl 中培养 只在14NH4Cl 中培养 提取DNA,进行密度梯度超离心 14N对照系统 15N对照系统 第一代 第二代
实验结果表明DNA复制是半保留复制 用15N标记的亲本DNA 14N中第一次复制 14N中第二次复制
Meselson-stahl 实验 (a)密度梯度离心的DNA带 (b)对应于左侧DNA带的解释
从前面图可以看出,在含有14N氮源介质中培养一代的DNA经密度梯度离心后,DNA带位于在15N培养基中的DNA带的上面。从前面图可以看出,在含有14N氮源介质中培养一代的DNA经密度梯度离心后,DNA带位于在15N培养基中的DNA带的上面。 在含有14N氮源介质中培养两代后的DNA经密度梯度离心后,出现两条带,第一条带位置与培养一带的DNA带相同,另一条DNA带位于第一条带上面,说明第二条带密度更轻。 实验结果充分说明DNA复制是半保留复制。
34.2 DNA复制是双向进行的 两种复制模式 (a)单向复制模式 (b)双向复制模式
复制起点 大肠杆菌染色体DNA双向复制模式 复制叉 复制叉
真核生物DNA复制有多个复制起点,同时双向复制真核生物DNA复制有多个复制起点,同时双向复制
34.3 DNA聚合酶III催化复制叉处的聚合反应 DNA合成时,核苷酸是通过酶催化加到一个正在延伸的DNA链上,这种酶要求一条完整的DNA链作为模板,去合成一条互补链,所以这样的酶叫做DNA指导的DNA聚合酶。 Arthur Kornberg(斯坦福大学医学院教授)等人从1955年开始寻找合成DNA的酶。于1956年在大肠杆菌的提取液中发现了DNA聚合酶。 Kornberg发现的DNA聚合酶就是现在的DNA聚合酶I,是分子量为109,000的一条多肽链,具有聚合活性及3ˊ-5ˊ外切酶活性,还有5ˊ-3ˊ核酸酶活性。以后又陆续发现了功能不同的另外几种DNA聚合酶,下表归纳了到目前为止从大肠杆菌和哺乳动物中发现的DNA聚合酶的基本特征。
聚合反应的基本过程都是通过新合成的链的3ˊ-OH对进入的新的核苷三磷酸(用d NTP)的磷进行亲核攻击,导致磷酯键断裂,结果在链的3ˊ末端加上了一个新的核苷酸,即延长了一个核苷酸。 释放出的焦磷酸经焦磷酸酶水解有利于聚合反应的进行
34.4 DNA聚合酶III同时催化两条链的合成 由于DNA的两条模板链是反平行的,又因为DNA总是沿5ˊ→ 3ˊ方向合成,即两条新链是沿着相反方向合成。换句话说,就是一条子链的合成方向与复制叉移动的方向一致,该子链称为前导链;而另一条子链与复制叉移动的方向相反,这条链称之为滞后链,但也是通过通过5ˊ→ 3ˊ方向聚合形成的。 9.4.1 在滞后链中DNA的合成是不连续的 根据岗崎实验,可以确定复制叉的一个“杈”(前导链)是沿5ˊ-3ˊ方向连续合成的,而另一条链(滞后链)是通过岗崎片段方式不连续合成的。连续和不连续合成的结合使得复制叉整体向一个方向延伸。
DNA聚合酶III同时催化两条链的合成 先导链 复制叉移动方向 岗崎片段 滞后链
34.4.2 每个岗崎片段的合成开始于一个RNA引物 不连续合成解释了滞后链是怎样合成的,但不能解释每个岗崎片段是怎样开始合成的。DNA聚合酶III不能从头开始进行聚合反应,它只能将核苷酸加到已有的多核苷酸链上。所以为了合成滞后链,需要在复制叉处合成一系列的短的RNA引物。每个RNA引物都是与滞后链模板部分互补的,而且在DNA聚合酶III催化下从引物 3ˊ端延伸形成岗崎片段。 RNA引物是通过引发酶合成的,引发酶是一个称为引发体的大的复合物中的成分,引发体还包括使DNA解旋的解旋酶和其它的一些辅助蛋白。引发酶每秒钟催化一个大约长10个核苷酸的RNA引物的合成。由于复制叉移动的速度大约为每秒1000个核苷酸,所以大约每1000个核苷酸就要合成一个RNA引物。
34.4.3 DNA pol I切去RNA引物,并使岗崎片段延伸 DNA pol I识别并结合于新DNA链的3ˊ端和下一个RNA引物之间的切口。然后5ˊ→ 3ˊ外切酶活性催化第1个RNA核苷酸水解,而5ˊ→ 3ˊ聚合酶活性将一个脱氧核苷酸加到新DNA链的3ˊ端,这种同时降解和合成的过程称为切口平移。 通过这种方式,使得切口沿滞后链移动。在完成切去RNA引物和填充RNA引物切去后的空缺后,DNA pol I 脱离DNA,留下了被一个切口(一个磷酸二酯键的空)分开的双链DNA。 最后在DNA连接酶催化下,一个岗崎片段的3ˊ末端和另一个岗崎片段的5ˊ磷酸基团之间形成一个磷酸二酯键,完成了两个岗崎片段的合成。
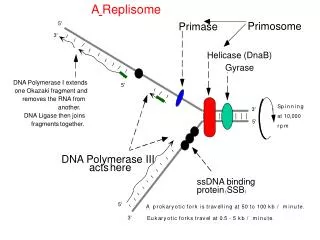
解旋酶 引发体 单链结合蛋白 引物酶 DNA聚合酶III DNA聚合酶I DNA连接酶 前导链 滞后链
34.5 复制叉移动需要多蛋白复合体 复制叉的移动除了需要用于聚合反应的DNA pol III以外,至少还需要四种其它蛋白质。其中主要的蛋白是解旋酶、拓扑异构酶、单链结合蛋白和DNA引发酶(也称为引物合成酶)。
解旋酶:是个需要ATP的酶,该酶刚好在移动的复制叉的前面沿着单链DNA滑动。E.coli中最重要的解旋酶是DnaB,它是一个与引发体中的引发酶紧密聚合的蛋白质。DnaB使复制叉前面的双螺旋DNA解旋,解旋过程是与核苷三磷酸水解过程相偶联的。解旋酶:是个需要ATP的酶,该酶刚好在移动的复制叉的前面沿着单链DNA滑动。E.coli中最重要的解旋酶是DnaB,它是一个与引发体中的引发酶紧密聚合的蛋白质。DnaB使复制叉前面的双螺旋DNA解旋,解旋过程是与核苷三磷酸水解过程相偶联的。 拓扑异构酶I和II:是用来将解旋酶作用后产生的扭转张力释放掉。DNA快速解旋在复制叉处形成超螺旋头部。拓扑异构酶I是切断DNA中的一条链,使DNA旋转,而拓扑异构酶II可以切断DNA的两条链。
单链结合蛋白(SSB):其作用是当解旋酶作用后,它可以防止解开的螺旋恢复原来状态。它对单链DNA有非常高的亲和性,在滞后链合成的开始需要单链结合蛋白,螺旋解旋后,前导链可以连续合成时就不需要这种蛋白了。单链结合蛋白(SSB):其作用是当解旋酶作用后,它可以防止解开的螺旋恢复原来状态。它对单链DNA有非常高的亲和性,在滞后链合成的开始需要单链结合蛋白,螺旋解旋后,前导链可以连续合成时就不需要这种蛋白了。 DNA引发酶:这个酶沿着滞后链在有规则的间隔内合成短的RNA引物。然后通过DNA pol III在引物的3ˊ-OH端沿着5ˊ-3ˊ方向合成岗崎片段。RNA引物可以被DNA pol I除去,然后用互补的脱氧核苷酸填上。在复制起点处前导链合成开始时也需要引发酶。
34.6 细菌中的DNA复制起始于染色体上唯一的一个部位 E.coli中,是位于称为oriC遗传位点的单一一个起点。oriC区含有两种重复类型的多个拷贝,DnaA结合部位含有一个特殊的9碱基对重复序列(4个拷贝),另一部位是富含A-T碱基对的重复区(3个拷贝)(图a)。 图b给出了在oriC处复制起始模式。大约有20个 DnaA蛋白与9碱基对重复区结合,并引起DNA结构的变化,导致富含13个A-T碱基对重复区内双螺旋变性。 富含A-T重复区的部位起着使复制泡中心定位的作用。一旦这个区接近其它DNA复制蛋白,在解旋酶和SSB作用下复制泡沿双向延伸形成两个复制叉。然后引发酶合成RNA引物,而DNA pol III开始前导链和滞后链的合成。
E.coli 的oriC结构模式:a. OriC内富含A-T序列和4个9碱基对重复序列的分布;b.在oriC处复制起始模式
34.7 DNA复制终止于ter区 在ter区内存在5个DNA序列(terA到terE)。ter序列排列在染色体上制造了一个复制叉“ 陷井”区,复制叉可以进入但不能出来。顺时针陷井由terB和terC构成,反时针陷井由terA、terD和terE构成。 陷井区是终止子利用物质(Tus)的结合部位。Tus可以和每一个ter结合。一旦形成Tus-ter复合物,就可以通过阻止解旋酶解旋DNA来封闭复制叉的通路。Tus-ter复合物只捕获来自一个方向(顺时针或反时针)的复制叉,而对来自另一个方向(反时针或顺时针)的复制叉不起作用。 终止区这样安排可确保两个从相反方向进入ter区的复制叉总能相遇,当一个复制叉遇到另一个复制叉时,DNA复制就完成了。
E.Coli中的ter区组织 600kb ter区含有5个非对称的ter部位,每个都可与Tus结合。 箭头表示为每个复制体设置的可能的终止部位
34.8 真核生物DNA复制类似于原核生物DNA复制 • 真核生物染色体一般都比细菌染色体大得多,如E,coli基因组是由4.6×103kb组成的单一染色体,而真核生物通常含有一个以上的染色体,一般在20至30对染色体之间。虽然染色体数目的增加使得基因组更复杂,但所有生物中的DNA复制的生物化学机制基本上是类似的。 • 真核生物DNA复制还有以下一些特点: • 真核生物所含的DNA聚合酶种类更多些。 • 真核生物复制叉处需要的辅助蛋白不同。 • 真核生物也象E.coli那样,复制是双向的。但E.coli染色体含有唯一的复制起点,而真核生物染色体存在许多复制起点。
34.9 DNA复制的其它方式 滚环复制 首先一个引发体组装在模板链(+链)上 ,开始合成RNA引物,然后在DNA pol III作用下,引物延伸生成一个互补链(-链)。 生成的双链DNA分子通过一个噬菌体编码的内切酶在+链上的特定部位制造一个缺口。 最后在DNA pol III作用下,+链从暴露出的3ˊ羟基延伸,取代原来的+链。
D-环复制 线粒体DNA的复制采取的是一种特殊的D-环复制方式: 环状双螺旋DNA在某一点解旋,开始复制。但前导链和滞后链的起点不在同一位点。 首先合成前导链,结果一条链(滞后链模板)仍维持单链。形成一个D字型的环。当前导链合成到某一点时,露出滞后链的起点,滞后链开始复制。
34.10 利用RNA作为模板,逆转录酶可以催化DNA合成 逆转录病毒的基因组是RNA分子,在感染期间,RNA分子可以逆转录为DNA分子。DNA再转录生产病毒RNA,或者与宿主DNA分子整合,使病毒潜伏于宿主后代中。 将逆转录病毒RNA转换成DNA的最关键酶是逆转录酶。该酶具有RNase H活性(降解RNA活性)和DNA聚合酶活性。
mRNA RNA-DNA杂化体 单链DNA 双链DNA mRNA的cDNA拷贝
① 逆转录病毒进入宿主细胞; ② 在逆转录酶的催化下,以病毒的RNA为模板合成互 补DNA(cDNA),形成RNA-DNA杂化体,逆转录酶 将杂化体中的RNA降解,同时以剩下的DNA链为模 板合成另一条互补的DNA链,结果形成双链DNA; ③ 双链DNA整合到宿主DNA中; ④ 利用宿主复制和转录机器生产大量的病毒RNA; ⑤ 转录出的mRNA翻译成病毒的包膜蛋白,逆转录酶 和壳体蛋白;
⑥ 将病毒RNA、逆转录酶和壳体蛋白组装成病毒 的核壳体; ⑦ 核壳体结合包膜蛋白形成完整的逆转录病毒。 逆转录酶没有3ˊ-5ˊ外切酶活性或校正活性,所以它的错误率比任何DNA聚合酶都高,例如人免疫缺陷病毒I(HIV I)逆转录酶每合成2000到4000核苷酸就会掺入一个不正确的碱基,这也部分说明了HIV I的高突变率。
34.11 损伤的DNA可以修复 1、脱嘌呤、脱氨和形成胸腺嘧啶二聚体都可能造成DNA损伤 DNA损伤常见形式是通过N-糖苷键的水解,鸟嘌呤或腺嘌呤的自发脱嘌呤作用形成脱氧核糖。据估计人体内细胞中每天每109碱基对就有多达103碱基对发生脱嘌呤。另一种类型损伤是胞嘧啶脱氨形成尿嘧啶,如果不能校正,在DNA复制后,一个C-G碱基对就将变成一个A-T碱基对了。
U C 胞嘧啶脱氨如不校正将导致: G-C突变为A-T U A
紫外线和离子辐射诱导有可能形成胸腺嘧啶二聚体。一个胸腺嘧啶的C-5、C-6与相邻胸腺嘧啶上同样位置的碳之间形成一个环丁基环,结果使得DNA骨架的结构发生了改变,有可能引起与互补链上的相应的腺嘌呤残基之间的氢键断裂。紫外线和离子辐射诱导有可能形成胸腺嘧啶二聚体。一个胸腺嘧啶的C-5、C-6与相邻胸腺嘧啶上同样位置的碳之间形成一个环丁基环,结果使得DNA骨架的结构发生了改变,有可能引起与互补链上的相应的腺嘌呤残基之间的氢键断裂。 胸腺嘧啶形成的示意图
2 在E.coli中存在4种基本的修复系统 E.coli中存在的4种基本类型的DNA修复系统:直接修复,核苷酸切除修复、碱基切除修复和错配修复。 A.直接修复 某些损伤的核苷酸和错配的碱基可以被某些蛋白质识别和修复。他们不切断DNA或切除碱基而是直接实施修复,这样的损伤修复机制称为直接修复。 DNA损伤之一的胸腺嘧啶二聚体可以通过直接修复机制修复。胸腺嘧啶二聚体是紫外线辐射造成的,光激活酶可以结合胸腺嘧啶二聚体引起的扭曲了的双螺旋部位。在可见光存在下,光激活酶催化两个胸腺嘧啶碱基再生,正常的A-T碱基对重新形成,然后光复活酶从已修复好的DNA上脱落。
B. 核苷酸切除修复 修复酶是由基因uvrA、uvrB和uvrC分别编码的三个亚基组成的,所以该酶又称为ABC切除核酸酶。 首先ABC切除核酸酶从损伤部位的两侧切去含有损伤的DNA链,结果都出现一个单链缺口。然后在DNA聚合酶催化下按照互补链填充缺口,切口最后通过DNA连接酶连接。
C. 碱基切除修复 DNA糖基化酶是一个能识别DNA中象尿嘧啶、次黄嘌呤和黄嘌呤那样的不正确碱基的酶,这些碱基都是由胞嘧啶、腺嘌呤和鸟嘌呤分别脱氨形成的。这样变型的碱基可以通过DNA糖基化酶切断N-糖苷键除去,这样一来在DNA中制造了脱嘌呤或脱嘧啶的部位,通常将这样的部位称之AP位点。 每种DNA糖基化酶通常对一种类型的碱基损伤特异。例如尿嘧啶糖基化酶就可以除去由于胞嘧啶脱氨形成的尿嘧啶,形成一个AP位点。然后一个称为AP内切核酸酶切去含有AP位点的脱氧核糖-5-磷酸,出现一个核苷酸空隙。然后在DNA聚合酶作用下重新放置一个正确的核苷酸,最后通过DNA连接酶将切口封闭。
D. 错配修复 偶尔错误的DNA复制会导致新合成的链与模板链之间的一个错误的碱基配对。这样的错误可以通过E.coli中的3个蛋白质(MutS、MutH和MutL)校正。该修复系统只能校正新合成的DNA,其主要依据是新合成的链中GATC序列中的A(腺苷酸残基)开始未被甲基化。GATC中A甲基化与否常用来区别新合成的链(未甲基化)和模板链(甲基化)。这一区别很重要,因为修复酶需要识别两个核苷酸残基中的哪一个是错配的,否则如果将正确的核苷酸除去就会导致突变。未甲基化的GATC序列不需要紧靠着错配碱基,因为错配碱基与GATC序列之间的间隔的DNA序列可以被外切核酸酶切除,是从3ˊ还是从5ˊ方向切除取决于不正确碱基的相对位置。
要点归纳 1. DNA复制是半保留复制,半保留复制已经经Meselson-Stahl实验证实。在半保留复制方式中两条亲代链分开,分开后的每一条链都可作为模板用于合成互补的、反平行的子链。就是说双螺旋子链中有一条链来自亲代链,另一条链则是以该亲代链为模板新合成的互补链。 2. DNA合成需要DNA聚合酶,DNA聚合酶需要一个游离的3′羟基作为引物(可以由RNA或DNA提供),以脱氧核苷三磷酸作为底物,催化 3′羟基与脱氧核苷三磷酸的5′-α-磷酸基团形成磷酸二酯键,释放出焦磷酸(随后水解为无机磷酸),使核苷酸整合到延伸的聚核苷酸链中。新链按5′→3′方向生长。
3.原核生物中存在3种聚合酶(pol I、II和III)。而真核生物中存在5种聚合酶(polα、β、γ、δ和ε)。在E.coli中,pol Ⅲ是主要的复制酶,而pol I既有复制功能,又有修复功能。所有三种原核生物的DNA聚合酶都具有3′→5′外切酶的活性,该酶可以将经聚合酶催化错配的核苷酸切去。DNA pol I和III还具有5′→3′外切酶活性。 4. 复制起始发生在复制起点,E.coli中存在唯一的复制起点(OriC),从一个复制起点沿着相反方向双向进行。真核生物染色体含有许多复制起点。真核生物DNA的复制也是双向进行的,但与E.coli不同,可以在许多复制起点同时双向进行复制。
5. DNA复制需要双链解旋形成复制叉,解旋需要解旋酶,ATP驱动的解旋酶使复制起点(OriC)区解旋,制造复制叉,单链结合蛋白与两条模板链结合防止他们重新形成双螺旋。在复制叉处,亲代DNA的两条链作为模板用于合成新的DNA。 6. 由于DNA聚合酶催化的链的延伸是5′→3′方向,以及双螺旋DNA中的两条链是反平行的,因此聚合酶沿着复制叉处两条模板链移动的方向是相反的。结果造成一条链(前导链)的合成是连续的,合成方向与复制叉移动方向一致;另一条链(滞后链)的合成是不连续的,合成方向与复制叉移动方向相反。
7.复制起始是借助于引发酶合成的RNA引物开始的,前导链合成需要一个RNA引物,而滞后链需要多个RNA引物。在滞后链合成中,在DNA pol III催化下,由每个引物按照模板链合成互补短DNA片段,称之为岗崎片段。然后DNA pol Ⅰ的5′→3′外切酶活性将 RNA 引物切除,再通过其聚合酶活性催化冈崎片段之间空隙的核苷酸填充,最后DNA连接酶将新生的冈崎片段连接起来。 8. 复制的忠实性非常高,主要是由于DNA聚合酶具有3′→5′外切酶的活性。但在复制过程中由于碱基配对错误、以及碱基共价修饰、碱基缺失和插入都会产生突变,并造成DNA损伤。复制后的修复主要包括5种修复机制:直接修复、切除修复、错配修复等。