Download
1 / 1
10 likes | 139 Views
Long-term Dynamics of Living and Dead Biomass Pools in an Old-Growth Northern Hardwood Forest Kerry D. Woods, Bennington College, Bennington, VT. INTRODUCTION:

E N D
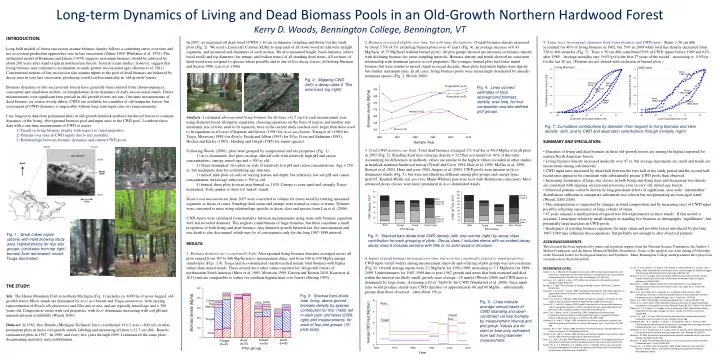
Long-term Dynamics of Living and Dead Biomass Pools in an Old-Growth Northern Hardwood Forest Kerry D. Woods, Bennington College, Bennington, VT INTRODUCTION: Long-held models of forest succession assume biomass density follows a saturating curve over time and net ecosystem production approaches zero in late succession (Odum 1969, Whittaker et al. 1974). The influential model of Bormann and Likens (1979) suggests maximum biomass should be achieved by about 200 years after stand origin in northeastern forests. Several recent studies, however, suggest that living biomass may continue to accumulate at much greater successional ages (Keeton et al. 2011). Conventional notions of late succession also assume inputs to the pool of dead biomass are balanced by decay rates in very late succession, producing overall carbon neutrality in 'old-growth' forests. Biomass dynamics in late-successional forests have generally been inferred from chronosequences, conceptual and simulation models, or extrapolations from dynamics of early-successional stands. Direct measurements over significant time periods in old-growth forests are rare. One-time measurements of dead biomass (or coarse woody debris, CWD) are available for a number of old temperate forests, but assessment of CWD dynamics is impossible without long-term input rates or remeasurements. I use long-term data from permanent plots in old-growth hemlock-northern hardwood forests to estimate dynamics of the living, aboveground biomass pool and input rates to the CWD pool. I combine these data with a one-time measurement of CWD to assess: 1) Trends in living biomass density with respect to stand properties; 2) Patterns over time in CWD inputs due to tree mortality; 3) Relationships between biomass dynamics and current CWD pools. In 2007, we measured all dead wood (CWD) > 10 cm in diameter (standing and down) for the study plots (Fig. 2). We used a Lasercraft Contour XLRic to map ends of all down wood in relatively straight segments, and measured end-diameters of each section. We also measured height, basal diameter (above basal swell) and top diameter (for stumps and broken stems) of all standing dead stems. All sections of dead wood were assigned to species where possible and to one of five decay classes (following Harmon and Sexton 1996, Liu et al. 2006). 2. Biomass increased slightly over time, but with large fluctuations: Overall biomass density increased by about 5.5% (4.5% excluding burned plots) over 47 years (Fig. 4), an average increase of 0.43 Mg/ha/yr (0.35 Mg/ha/yr without burned plots). All plot groups showed net increases in biomass density with declining biomass for some sampling intervals. Biomass density and trends showed no consistent relationship with dominant species or soil properties. The younger, burned plots had lower initial biomass but were similar to mixed stands in recent decades; these plots had much higher stem density but smaller maximum sizes. In all cases, living biomass pools were increasingly dominated by already-dominant species (Fig. 2, Woods 2000). 5: Large trees increasingly dominate both living biomass and CWD input :Stems > 50 cm dbh accounted for 40% of living biomass in 1962, but 59% in 2009 while total tree density decreased from 520 to 406 stems/ha (Fig. 7). Trees > 50 cm dbh contributed 50% of CWD inputs before 1989 and 62% after 1989. Average mortality rate 0.6%/yr for the first 27 years of the record, , increasing to 0.9%/yr for the last 20 yrs. (Patterns are not altered with exclusion of burned plots.) Fig. 2: Mapping CWD (left); a decay-class 5 Tiliaamericanalog (right). Fig. 4: Lines connect estimates of total, aboveground biomass density, over time, for four composition and site-defined plot groups. Analysis: I estimatedaboveground living biomass for all trees >12.5 cm for each measurement year, using diameter-based allometric equations, choosing equations on the basis of region, and number and maximum size of trees used to fit equation; trees in the current study reached sizes larger than those used to fit equations in all cases (Chapman and Gower (1991) for Acer saccharum, Young et al. (1980) for Tsuga, Morrison (1990) for Betula, Perala and Alban (1993)for Tilia, Crow and Erdmann (1983), Hocker and Earley (1983), Harding and Grigal (1985) for minor species). Following Woods (2000), plots were grouped by composition and site properties (Fig. 1): 1) Acer-dominated: five plots on deep, alluvial soils with relatively high pH and cation concentrations; canopy mixed-age and > 300 yr old. 2) Tsuga-dominated: two plots on soils of relatively low pH and cation concentrations. Age > 250 yr, but inadequate data for establishing age structure. 3) mixed: nine plots on soils of varying texture and depth, but relatively low soil pH and cation concentration; mixed-age with trees > 300 yr old 4) burned: three plots from an area burned ca. 1830. Canopy is even-aged and strongly Tsuga-dominated. Soils similar to those for 'mixed' stands. Dead wood measurements from 2007 were converted to volume for down wood by treating measured segments as frusta of cones. Standing dead stems and stumps were treated as cones or frusta. Volumes were converted to mass using relationships specific to decay class and species from Liu et al. (2006). CWD inputs were calculated from mortality between measurements using stem-only biomass equations with last recorded diameter. This neglects contributions of large branches, but these constitute a small proportion of both living and dead biomass. Any diameter growth between last live measurement and tree death is also discounted, which may be of consequence only for the long 1967-1989 interval. Fig. 7: Cumulative contributions by diameter (from largest) to living biomass and stem density (left), and to CWD and dead stem contributions through mortality (right). • SUMMARY AND SPECULATION: • Densities of living and dead biomass in these old-growth forests are among the highest reported for eastern North American forests. • Living biomass density increased modestly over 47 yr, but average increments are small and trends are inconsistent among sub-intervals. • CWD input rates increased by about half between the first half of the study period and the second half; recent rates appear to be consistent with substantially greater CWD pools than observed. • Increasing dominance of large size classes in both living and dying trees and decreasing tree density are consistent with ongoing successional processes even in very old, mixed-age forests. • Observed patterns could be driven by long-persistent effects of significant, area-wide ‘intermediate’ disturbances sufficient to inaugurate substantial tree cohorts but not generating an even-aged stand (Woods 2000,2004). • This interpretation is supported by changes in stand composition and by increasing rates of CWD input possibly reflecting senescence of large cohorts of stems. • 47 years remains a small portion of typical tree life-expectancies in these stands. If this model is accurate, I anticipate relatively small changes in standing live biomass as demography ‘equilibrates’, but potentially large increases in CWD pools. • Inadequacy of existing biomass equations for large stems and possible biases introduced by the long 1967-1989 may influence these equations, but probably not enough to alter observed patterns. 3. Total CWD densities are high:Total dead biomass averaged 151.4 m3/ha or 46.0 Mg/ha over all plots in 2007 (Fig. 5). Standing dead trees (average density = 32.5/ha) accounted for 16% of this total. Accounting for differences in methods, values are similar to the highest values recorded in other studies in hemlock-northern hardwood forests (Tyrrell and Crow 1994, Hale et al. 1999, McGee et al. 1999, Stewart et al. 2003, Hura and crow 2004, Angers et al. 2005). CWD pools were greatest in Acer-dominated stands (Fig. 5), but were not otherwise different among plot groups and canopy types (p>0.05, Kruskal-Wallis test, post hoc Mann-Whitney pair-wise tests with Bonferroni correction). Most advanced decay classes were more prominent in Acer-dominated stands. Fig. 1: Great Lakes region (above) with inset showing study area. Habitat photos for four plot groups: (clockwise from top right) burned; Acer-dominated; mixed; Tsuga-dominated. Fig. 5: Stacked bars divide total CWD density (left) and volume (right) by decay class contribution for each grouping of plots. Decay class 1 includes stems with no evident decay; decay class 5 includes sections with little or no solid wood or structure. ACKNOWLEDGEMENTS: This research has been supported by grants and logistical support from the National Science Foundation, the Andrew J. Mellon Foundation, and the Huron Mountain Wildlife Foundation. Some of the analysis was done during a Fellowship at the National Center for Ecological Analysis and Synthesis. Many Bennington College undergraduates have played an essential role in the field and lab. RESULTS: 1. Biomass densities are exceptionally high:Aboveground living biomass densities averaged across all plots ranged from 367 to 406 Mg/ha across measurement dates, and from 340 to 430 Mg/ha among stand-types (Figs. 3,4). Tsuga and Acer-dominated stands reached similar total biomass with higher values than mixed stands. These exceed most other values reported for old-growth forests of northeastern North America (Mroz et al. 1985, Morrison 1990, Curzon and Keeton 2010, Keeton et al. 2011) and are comparable to values for southern Appalachian cove forests (Busing 1993). 4. Inputs of dead biomass increased over time, but were not consistently related to stand properties: CWD input varied widely among measurement intervals and ordering of plot groups was not consistent (Fig. 6). Overall average inputs were 2.1 Mg/ha/yr for 1962-1989, increasing to 3.1 Mg/ha/yr for 1989-2009. Underestimates for 1967-1989 due to post-1967 growth and stems that both recruited and died within the interval are likely small; growth rates average ~20 mm/yr (Woods 2000) and CWD mass is dominated by large stems. Assuming a 20-yr ‘half-life’ for CWD (Vanderwel et al. 2008), these input rates would produce steady-state CWD densities of approximately 60 and 90 Mg/ha – substantially greater than those observed – after about 150 yr. REFERENCES CITED: Angers, V. A., C. Messier, M. Beaudet, and A. Leduc. 2005. Comparing composition and structure in old-growth and harvested (selection and diameter-limit cuts) northern hardwood stands in Quebec. For. Ecol. and Manage. 217:275–293. Bormann, F. H., and G. E. Likens. 1979. Pattern and process in a forested ecosystem. Springer-Verlag, New York. Busing, R. T. 1993. Three Decades of Change at Albright Grove, Tennessee. Castanea 58:231–242. Chapman, J. W., and S. T. Gower. 1991. Aboveground production and canopy dynamics in sugar maple and red oak trees in southwestern Wisconsin. Can. J. Forest Res. 21:1533–1543.. Crow, T. R., and G. G. Erdmann. 1983. Weight and volume equations and tables for red maple in the Lake States. Research Paper NC-242. USDA Forest Service. Curzon, M. T., and W. S. Keeton. 2010. Spatial characteristics of canopy disturbances in riparian old-growth hemlock-northern hardwood forests, Adirondack Mountains, New York, USA. Canadian Journal of Forest Research 40:13–25. Hale, C. M., J. Pastor, and K. A. Rusterholz. 1999. Comparison of structural and compositional characteristics in old-growth and mature, managed hardwood forests of Minnesota, USA Can. J. Forest Res. 29:1479–1489. Harding, R. B., and D. F. Grigal. 1985. Individual tree biomass estimation equations for plantation-grown white spruce in northern Minnesota. Can. J. Forest Res. 15:738–739. Harmon, M. E., and J. Sexton. 1996. Guidelines for measurements of woody detritus in forest ecosystems. US LTER Network Office. Hocker, H. W., and D. J. Earley. 1983. Biomass and leaf area equations for northern forest species. New Hampshire Agricultural Experiment Station. Hura, C. E., and T. R. Crow. 2004. Woody debris as a component of ecological diversity in thinned and unthinned northern hardwood forests. Nat. Areas Journal 24:57–64. Keeton, W. S., A. A. Whitman, G. C. McGee, and C. L. Goodale. 2011. Late-successional biomass development in northern hardwood-conifer forests of the northeastern United States. Forest Science 57:489–505. Liu, W. H., D. M. Bryant, L. R. Hutyra, S. R. Saleska, E. Hammond-Pyle, D. Curran, and S. C. Wofsy. 2006. Woody debris contribution to the carbon budget of selectively logged and maturing mid-latitude forests. Oecologia 148:108–117. McGee, G. G., D. J. Leopold, and R. D. Nyland. 1999. Structural Characteristics of Old-Growth, Maturing, and Partially Cut Northern Hardwood Forests. Ecological Applications 9:1316–1329. Morrison, I. K. 1990. Organic matter and mineral distribution in an old-growth Acersaccharum forest near the northern limit of its range. Can. J. Forest Res. 20:1332–1342. Mroz, G. D., M. R. Gale, M. F. Jurgensen, D. J. Frederick, and A. Clark III. 1985. Composition, structure, and above ground biomass of two old-growth northern hardwood stands in Upper Michigan. Can. J. Forest Res. 15:78–82. Odum, E. P. 1969. The strategy of ecosystem development. Science 164:262–270. Perala, D. A., and D. Alban. 1993. Allometric biomass estimators for aspen-dominated ecosystems in the upper Great Lakes. Research Paper NC-314. USDA Forest Service Stewart, B. J., P. D. Neily, E. J. Quigley, and L. K. Benjamin. 2003. Selected Nova Scotia old-growth forests: Age, ecology, structure, scoring. The Forestry Chronicle 79:632–644. Tyrrell, L. E., and T. R. Crow. 1994. Structural characteristics of old-growth hemlock-hardwood forests in relation to age. Ecology 75:370–386. Vanderwel, M. C., H. C. Thorpe, J. L. Shuter, J. P. Caspersen, and S. C. Thomas. 2008. Contrasting downed woody debris dynamics in managed and unmanaged northern hardwood stands. Can. J. Forest Res. 38:2850–2861. Whittaker, R. H., F. H. Bormann, G. E. Likens, and T. G. Siccama. 1974. The Hubbard Brook ecosystem study: forest biomass and production. Ecol. Monogr 44:233–254. Woods, K. D. 2000. Dynamics in late-successional hemlock-hardwood forests over three decades. Ecology 81:110–126. Woods, K. D. 2004. Intermediate disturbance in a late-successional hemlock-northern hardwood forest. J. Ecol. 92:464–476. Young, H. E., J. H. Ribe, and K. Wainwright. 1980. Weight tables for tree and shrub species in Maine. Maine. Life Sciences and Agriculture Experiment Station. Miscellaneous report. THE STUDY: Site: The Huron Mountain Club in northern Michigan (Fig. 1) includes ca. 4000 ha of never-logged, old-growth forest. Mesic stands are dominated by Acer saccharum and Tsugacanadensis, with varying representation of BetulaalleghaniensisandTiliaamericana, and are typically mixed-age and > 300 years old. Composition varies with soil properties, with Acer dominance increasing with soil pH and mineral nutrient availability (Woods 2000). Data-set:In 1962, Eric Bourdo (Michigan Technical Univ.) established 19 0.2 acre (~800 m2) circular permanent plots in mesic old-growth stands, labeling and measuring all trees > 12.5 cm dbh. Bourdoremeasured plots in 1967. In 1989, and every five years through 2009, I remeasured the same plots, documenting mortality and establishment. Fig. 3: Stacked bars divide total living, above-ground biomass density by species contribution for first (1962, left in each pair) and latest (2009, right) plot measurements, for each of four plot groups (19 plots total). Fig. 6: Lines indicate average annual inputs of CWD (standing and down combined) via tree mortality by measurement interval and plot group. Values are for stem or bole only, estimated from last living diameter measurement.