Download
1 / 29
290 likes | 345 Views
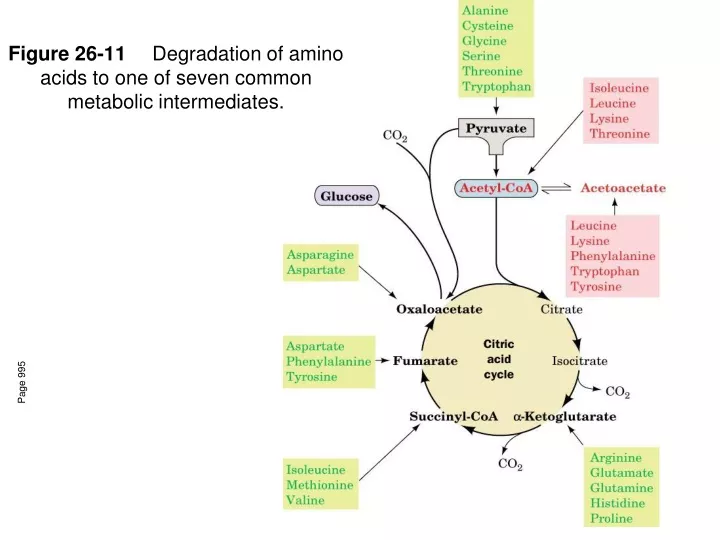
Figure 26-11 Degradation of amino acids to one of seven common metabolic intermediates. Page 995. Met degradation. Met reacts with ATP to form S- adenosylmethionine (SAM). SAM’s sulfonium ion is a highly reactive methyl group so this compound is involved in methylation reactions.

E N D
Figure 26-11 Degradation of amino acids to one of seven common metabolic intermediates. Page 995
Met degradation • Met reacts with ATP to form S-adenosylmethionine (SAM). • SAM’s sulfonium ion is a highly reactive methyl group so this compound is involved in methylation reactions. • Methylation reactions catalyzed by SAM yield S-adenosylhomocysteine and a methylated acceptor molecule. • S-adenosylhomocysteine is hydrolyzed to homocysteine. • Homocysteine may be methylated to regenerate Met, in a B12 requiring reaction with N5-methyl-THF as the methyl donor. • Homocysteine can also combine with Ser to form cystathionine in a PLP catalyzed reaction and -ketobutyrate. • -ketobutyrate is oxidized and CO2 is released to yield propionyl-CoA. • Propionyl-CoA proceeds thorugh to succinyl-CoA.
Methionine adenosyltransferase Methyltransferase Adenosylhomocysteinase Methionine synthase (B12) Cystathionine -synthase (PLP) Cystathionine -synthase (PLP) -ketoacid dehydrogenase Propionyl-CoA carboxylase (biotin) Methylmalonyl-CoA racemase Methylmalonyl-CoA mutase Glycine cleavage system or serine hydroxymethyltransferase N5,N10-methylene-tetrahydrofolate reductase (coenzyme B12 and FAD) NADH, H+ Page 1002
Branched chain amino acid degradation • Degradation of Ile, Leu, and Val use common enzymes for the first three steps • Transamination to the corresponding -keto acid • Oxidative decarboxylation to the corresponding acyl-CoA • Dehydrogenation by FAD to form a double bond. First three enzymes • Branched-chain amino acid aminotransferase • Branched-chain keto acid dehydrogenase (BCKDH) • Acyl-CoA dehydrogeanse
Figure 26-21 The degradation of the branched-chain amino acids (A) isoleucine, (B) valine, and (C) leucine. Page 1004
After the three steps, for Ile, the pathway continues similar to fatty acid oxidation (propionyl-CoA carboxylase, methylmalonyl-CoA racemase, methylmalonyl-CoA mutase). • Enoyl-CoA hydratase - double bond hydration • -hydroxyacyl-CoA dehydrogenase- dehydrognation by NAD+ • Acetyl-CoA acetyltransferase - thiolytic cleavage Page 1004
For Valine: • Enoyl-CoA hydratase - double bond hydration • -hydroxy-isobutyryl-CoA hydrolase -hydrolysis of CoA • hydroxyisobutyrate dehydrogenase - second dehydration • Methylmalonate semialdehyde dehydrogenase - oxidative carboxylation • Last 3 steps similar to fatty acid oxidation Page 1004
For Leucine: • -methylcronyl-CoA carboxylase-carboxylation reaction (biotin) • -methylglutaconyl-CoA hydratase-hydration reaction • HMG-CoA lyase • Acetoacetate can be converted to 2 acetyl-CoA • Leucine is a ketogenic amino acid! Page 1004
Leu and Lys are ketogenic • Leu proceeds through a typical branched amino acid breakdown but the final products are acetyl-CoA and acetoacetate. • Most common Lys degradative pathway in liver goes through the formation of the -ketoglutarate-lysine adduct saccharopine. • 7 of 11 reactions are found in other pathways. • Reaction 4: PLP-dependent transamination • Reaction 5: oxidative decarboxylation of an a-keto acid by a multienzyme complex similar to pyruvate dehydragense and a-ketoglutarate dehydrogenase. • Reactions 6,8,9: fatty acyl-CoA oxidation. • Reactions 10 and 11 are standard ketone body formation reactions.
Figure 26-23 The pathway of lysine degradation in mammalian liver. Saccharopine dehydrogenase (NADP+, Lys forming) Saccharopine dehydrogenase (NAD+, Glu forming) Aminoadipate semialdehyde dehydrogenase Aminoadipate aminotransferase (PLP) -keto acid dehydrogenase Glutaryl-CoA dehydrogenase Decarboxylase Enoyl-CoA hydratase -hydroxyacyl-CoA dehydrogenase HMG-CoA synthase HMG-CoA lyase Page 1006
Trp is both glucogenic and ketogenic • Trp is broken down into Ala (pyruvate) and acetoacetate. • First 4 reactions lead to Ala and 3-hydroxyanthranilate. • Reactions 5-9 convert 3-hydroxyanthranilate to a-ketoadipate. • Reactions 10-16 are catalyzed by enzymes of reactions 5 - 11 in Lys degradation to yield acetoacetate.
1. Tryptophan-2,3-dioxygenase, 2. Formamidase, 3. Kynurenine-3-monooxygense, 4. kynureninase (PLP dependent) Page 1007
Kynureinase, another PLP mechanism • Reaction 4: cleavage of 3-hydroxykynurenine to alanine and 3-hydroxyanthranilate is catalyzed by the PLP dependent enzyme kynureinase. • This facilitates a C-C bond cleavage. (previous reactions catalyzed the C-H and C-C bond cleavage) • Follows the same steps as transamination but does not hydrolyze the tautomerized Schiff base. • Enzyme amino acid acts as a nucleophile tto attack the carbonyl carbon (Cof the tautomerized 3-hydroxykynurenine-PLP Schiff base.
6. Amino carboxymuiconate semialdehyde decarboxylase 7. Aminomuconate semialdehyde dehydrogenase 8. Hydratase, 9. Dehydrogense 10-16. Reactions 5-11 in lysine degradation. Page 1007
-keto acid dehydrogenase • Glutaryl-CoA dehydrogenase • Decarboxylase • Enoyl-CoA hydratase • -hydroxyacyl-CoA dehydrogenase • HMG-CoA synthase • HMG-CoA lyase Page 1006
Phe and Tyr are degraded to fumarate and acetoacetate • The first step in Phe degradation is conversion to Tyr so both amino acids are degraded by the same pathway. • 6 reactions
Phenylanalnine hydroxylase • Aminotransferase • p-hydroxyphenylpyruvate dioxygenase • Homogentisate dioxygenase • Maleylacetoacetate isomerase • Fumarylacetoacetase Page 1009
Phenylalanine hydroxylase has biopterin cofactor • 1st reaction is a hydroxylation reaction by phenylalanine hydroxylase (PAH), a non-heme-iron containing homotetrameric enzyme. • Requires O2, FeII, and biopterin a pterin derivative. • Pterins have a pteridine ring (similar to flavins) • Folate derivatives (THF) also contain pterin rings.
Figure 26-27 The pteridine ring, the nucleus of biopterin and folate. Page 1009
Active BH4 must be regenerated • Active form in PAH is 5,6,7,8-tetrahydrobiopterin (BH4) • Produced from 7,8-dihydrobiopterin via dihydrofolate reductase (NADPH dependent). • 5,6,7,8-tetrahydrobiopterin is hydroxylated to pterin-4a-cabinolamine by phenylalanine hydroxylase. • pterin-4a-cabinolamine is converted to 7,8-dihydrobiopterin (quinoid form) by pterin-4a-carbinoline dehydratase • 7,8-dihydrobiopterin (quinoid form) is reduced by dihydropteridine reductase to regenerate the active cofactor.
NIH shift • A 3H that starts on C4 of Phe’s ring ends up on C3 of Tyr’s ring rather than being lost to solvent. • Mechanism is called the NIH shift • 1st characterized by scientists at NIH
1 and 2: activation of the enzyme’s BH4 and Fe(II) cofactors to yield pterin-4a-carbinolamine and a reactive oxyferryl [Fe(IV)=O2-] 3: Fe(IV)=O2- reacts with Phe to form an epoxide across the 3,4 bond. 4: epoxide opening to form carbocation at C3 5: migration of hydride from C4 to C3 to form more stable carbocation. 6: ring aromatization to form Tyr
Phe and Tyr are degraded to fumarate and acetoacetate • The first step in Phe degradation is conversion to Tyr so both amino acids are degraded by the same pathway. • 6 reactions • Reaction 1 = 1st NIH shift • Reaction 3 is also an example of NIH shift (26-31)
Phenylanalnine hydroxylase • Aminotransferase • p-hydroxyphenylpyruvate dioxygenase • Homogentisate dioxygenase • Maleylacetoacetate isomerase • Fumarylacetoacetase Page 1009
Amino acids as precursors • Amino acids are essential precursors to biomolecules: • Nucleotides • Nucleotide coenzymes • Heme • Hormones • Neurotransmitters • Glutathione