Download

1 / 28
280 likes | 299 Views
Learn about how cells produce energy with and without oxygen through glycolysis, fermentation, and metabolic pathways in this comprehensive guide.

E N D
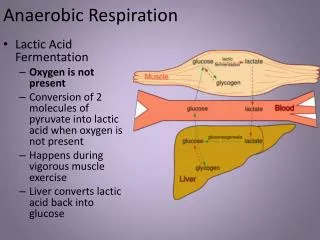
Under anaerobic conditions, the NADH cannot be reoxidized through the respiratory chain to oxygen. Pyruvate is reduced by the NADH to lactate,catalyzed by lactate dehydrogenase. There are different tissue specific isoenzymes lactate dehydrogenase . The reoxidation of NADH via lactate formation allows glycolysis to proceed in the absence of oxygen by regenerating sufficient NAD+ for another cycle of the reaction catalyzed by glyceraldehyde 3-phosphate dehydrogenase.
Fermentation: Anaerobic organisms lack a respiratory chain. They must reoxidize NADHproduced in Glycolysis through some other reaction, because NAD+ is needed for the Glyceraldehyde-3-phosphate Dehydrogenase reaction. Usually NADH is reoxidized as pyruvate is converted to a more reduced compound. The complete pathway, including Glycolysis and the reoxidation of NADH, is called fermentation.
E.g., Lactate Dehydrogenase catalyzes reduction of the keto in pyruvate to a hydroxyl, yielding lactate, as NADH is oxidized to NAD+. Cell membranes contain carrier proteins that facilitate transport of lactate.
Skeletal muscles ferment glucose to lactate during exercise, when the exertion is brief and intense. Lactate released to the blood may be taken up by other tissues, or by skeletal muscle after exercise, and converted via Lactate Dehydrogenase back to pyruvate, which may be oxidized in Krebs Cycle or (in liver) converted to back to glucose via gluconeogenesis
Some anaerobic organisms metabolize pyruvate to ethanol, which is excreted as a waste product. NADH is converted to NAD+ in the reaction catalyzed by Alcohol Dehydrogenase.
Glycolysis: glucose + 2 NAD+ + 2 ADP + 2 Pi 2 pyruvate + 2 NADH + 2 ATP Fermentation, from glucose to lactate: glucose + 2 ADP + 2 Pi2 lactate + 2 ATP Anaerobic catabolism of glucose yields only 2 “high energy” bonds of ATP.
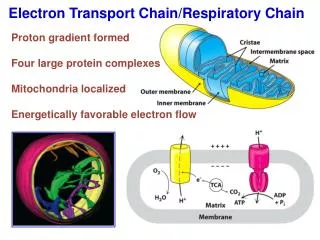
Under aerobic conditions, pyruvate is taken up into mitochondria,and after oxidative decarboxylation to acetyl-CoA is oxidized to CO2 by the citric acid cycle.
*Values in this table from D. Voet & J. G. Voet (2004) Biochemistry, 3rd Edition, John Wiley & Sons, New York, p. 613.
Hexokinase is inhibited by productglucose-6-phosphate: • by competition at the active site • by allosteric interaction at a separate enzyme site. Cells trap glucose by phosphorylating it, preventing exit on glucose carriers. Product inhibition of Hexokinase ensures that cells will not continue to accumulate glucose from the blood, if [glucose-6-phosphate] within the cell is ample.
Glucokinase, with high KM for glucose, allows liver to store glucose as glycogen in the fed state when blood [glucose] is high. Glucose-6-phosphatase catalyzes hydrolytic release of Pi from glucose-6-P. Thus glucose is released from the liver to the blood as needed to maintain blood [glucose]. The enzymes Glucokinase & Glucose-6-phosphatase, both found in liver but not in most other body cells, allow the liver to control blood [glucose].
Pyruvate Kinase, the last step Glycolysis, is controlled in liver partly by modulation of the amount of enzyme. This facilitates converting excess glucose to pyruvate, which is metabolized to acetyl-CoA, the main precursor for synthesis of fatty acids, for long term energy storage.
Phosphofructokinase is usually the rate-limiting step of the Glycolysis pathway. Phosphofructokinase is allosterically inhibited byATP. • At low concentration, the substrate ATP binds only at the active site. • At high concentration, ATP binds also at a low-affinity regulatory site, promoting the tense conformation.
Inhibition of the Glycolysis enzyme Phosphofructokinase when [ATP] is high prevents breakdown of glucose in a pathway whose main role is to make ATP. It is more useful to the cell to store glucose as glycogen when ATP is plentiful.
In Erythrocytes, the First Site of ATP Formation in Glycolysis May Be Bypassed In erythrocytes, the reaction catalyzed by phosphoglycerate kinase may be bypassed to some extent by the reaction of bisphosphoglycerate mutase, which catalyzes the conversion of 1,3-bisphosphoglycerate to 2,3- bisphosphoglycerate, followed by hydrolysis to 3-phosphoglycerate and Pi , catalyzed by 2,3- bisphosphoglycerate phosphatase. This alternative pathway involves no net yield of ATP from glycolysis. However, it does serve to
provide 2,3-bisphosphoglycerate, which binds to hemoglobin, decreasing its affinity for oxygen, and so making oxygen more readily available to tissues This is called as Rapaport-Luebering cycle.
Regulation of glycolesis • The three irreversible steps, catalyzed by hexokinase , phosphofructokinase and pyruvate kinase are the sites of regulation of glycolysis. Phosphofructokinaseis significantly inhibited at normal intracellular concentrations of ATP. • Insulin stimulates the activities of these enzymes thereby increasing the utilization of glucose in glycolysis. On the other hand, glucagon inhibits the process.
Metabolic Fates of Pyruvate • Under Anaerobic Condition Under anaerobic conditions, pyruvate is reduced to lactate. This reaction is catalyzed by lactate dehydrogenase and requires NADH+H+ .
Under Aerobic Condition Under aerobic condition, pyruvate is transported to the mitochondria where it is oxidativelydecarboxylated to acetyl CoA, by pyruvatedehydrogenase complex. This enzyme requires five coenzymes, TPP, CoA, NAD+ , FAD and Lipoic acid.
THE OXIDATION OF PYRUVATE TO ACETYL-COA IS THE IRREVERSIBLEROUTE FROM GLYCOLYSIS TO THE CITRIC ACID CYCLE
Pyruvate dehydrogenase Complex (PDC) • It is a multi-enzyme complex containing three enzymes associated together non-covalently: • E-1 : Pyruvate dehydrogenase , uses Thiamine pyrophosphate as cofactor bound to E1 • E-2 : Dihydrolipoyl transacetylase, Lipoic acid bound, CoA as substrate • E-3 : Dihydrolipoyl Dehydrogenase FAD bound, NAD+ as substrate • Advantages of multienzyme complex: • Higher rate of reaction • Minimum side reaction. • Coordinated control.
Pyruvate Dehydrogenase Is Regulated by End-Product Inhibition & Covalent Modification • Pyruvate dehydrogenase is inhibited by its products, acetyl-CoA and NADH .It is also regulated by phosphorylation by a kinase of three serine residues on the pyruvate dehydrogenase component of the multienzyme complex, resulting in decreased activity and by dephosphorylation by a phosphatase that causes an increase in activity.
The kinase is activated by increases in the [ATP]/[ADP], [acetyl-CoA]/[CoA], and [NADH]/[NAD+ ] ratios. Thus, pyruvate dehydrogenase, and therefore glycolysis, is inhibited both when there is adequate ATP (and reduced coenzymes for ATP formation) available, and also when fatty acids are being oxidized.
Energy Production During Glycolysis • Total number of ATP produced during glycolysis varies, depending upon conditions of the reactions and the concentration of substrates. Under Anaerobic Conditions Since each of the energy-yielding steps, i.e. the conversion of 1,3-bisphosphoglyceratee to 3-phosphoglycerate and phosphoenolpyrovate to pyrovate, produces one ATP, a total of 2 ATP are produced for each molecule of the triose oxidized or 4 molecules of ATP per molecule of hexose
As 2 ATP are used in the initial reactions of the process, i.e. in the conversion of glucose to glucose 6-phosphate and fructose-6-phosphate to fructose-1,6-bisphosphate,the net yield, per molecules of glucose, is only 2 ATP . • Under aerobic Conditions • In this process, NADH produced during the conversion of 3-phosphoglycerate to 1,3 bis –phosphoglycerate enters the electron transport chain and relases 3 ATP. Thus, additionally 6ATP are produced per molecule of glucose. The total yield therefore is 10 ATP and the net yield is 8 ATP.