Download

1 / 26
280 likes | 339 Views
Explore the structural levels of DNA in eukaryotic genomes, from a single linear DNA double helix to chromosome formation. Learn about chromatin fiber, regulation of chromatin structure, epigenetic inheritance, cell differentiation, and transcription regulation. Discover how gene expression is controlled through transcription initiation, control elements, and cell type-specific transcription. Delve into post-transcriptional regulation mechanisms like alternative RNA splicing, mRNA degradation, miRNAs, and RNA interference.

E N D
Eukaryotic Genomes CHAPTER 19
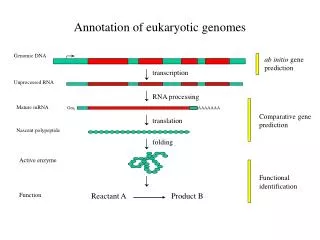
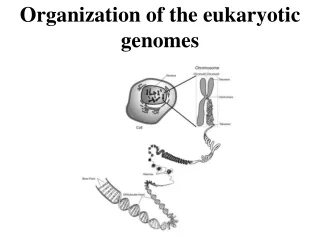
Structural Levels of DNA • a single linear DNA double helix averages about 4 cm in length • DNA associates with proteins that condense it so it will fit in the nucleus • DNA-protein complex = chromatin • chromatin looks like “beads on a string” when unfolded • “beads” = nucleosomes made up of histones (proteins) “string” = DNA • http://www.youtube.com/watch?v=9kQpYdCnU14&feature=relmfu • http://www.youtube.com/watch?v=gbSIBhFwQ4s&feature=relmfu
chromatin fiber (30 nm) • created by interactions between adjacent nucleosomes and the linker DNA • chromatin fiber (300 nm) • created when the 30 nm chromatin fiber forms loops called looped domains attached to a protein scaffold made of nonhistones • chromosome • forms when the 300 nm chromatin fiber folds on itself
Regulation of Chromatin Structure • compactness of chromatin helps regulate gene expression • heterochromatin – highly compact so it is inaccessible to transcription enzymes • euchromatin – less compact allowing transcription enzymes access to DNA • chemical modifications that can alter chromatin compactness: • histoneacetylation (-COCH3) neutralizes the histones so they no longer bind to neighboring nucleosomes causing chromatin to have a looser structure
DNA Methylation • addition of methyl groups to DNA bases (usually cytosine) inactivate DNA • methylation patterns can be passed on • after DNA replication, methylation enzymes correctly methylate the daughter strand • accounts for genomic imprinting in mammals – expression of either the maternal or paternal allele of certain genes during development • (NOTE: inheritance of chromatin modifications that do not involve a change in the DNA sequence is called epigenetic inheritance) • http://www.youtube.com/watch?NR=1&v=dfdnf1Wpg0E&feature=endscreen
Cell Differentiation • process of cell specialization (form & function) during the development of an organism • differences in cell types results from differential gene expression • several control points at which gene expression can be regulated (turned on/off, accelerated, slowed down) • most commonly regulated at transcription in response to an extracellular signal
Regulation of Transcription Initiation • general transcription factors – proteins that form a transcription initiation complex on the promoter sequence (ex: TATA box) allowing RNA polymerase to begin transcription • control elements – segments of noncoding DNA that help regulate transcription by binding certain proteins • proximal control elements • distal control elements (enhancers) - interact with specific transcription factors: • activators –stimulate transcription by binding to enhancers • repressors - inhibit transcription by binding directly to enhancers or by blocking activator binding to enhancers or other transcription machinery
activators bind to enhancer with 3-binding sites • a DNA-bending protein brings the bound activators closer to the promoter • activators bind to general transcription factors & mediator proteins, helping them to form a functional transcription initiation complex • activators can also promote histone acetylation & repressors can promote histone deacetylation
Cell Type-Specific Transcription • the # of different genes far exceeds the # of different control elements; therefore, the particular combination of control elements is what is important in controlling transcription along with the available activators in the cell
Co-expressed Genes • most co-expressed genes are found scattered over different chromosomes • to coordinate their gene expression, each gene is regulated by the same control elements & these control elements are activated by the same chemical signals
Post-Transcriptional Regulation • alternative RNA-splicing: regulatory proteins specific to a cell type control intron-exon choices, thereby producing different mRNA molecules from the same primary transcript
methods of mRNA degradation: • enzymatic shortening of the poly-A tail triggers the removal of the 5′ cap which is followed by the digestion of the mRNA by nucleases • nucleotide sequences in the untranslated region at the 3′ end regulate the length of time an mRNA remains intact • microRNAs (miRNAs) – small RNA molecules that bind to complementary sequences on mRNA causing degradation by associated proteins; miRNAs may also block translation • RNA interference (RNAi) pathway – involve small interfering RNAs (siRNAs) that function in the same way as miRNAs
initiation of translation: • can be blocked by regulatory proteins that bind to specific sequences or structures within the untranslated region at the 5’ end of the mRNA & prevent the attachment of ribosomes • translation will not begin if the poly-A tails are not long enough • “global” control – activation or inactivation of one or more of the protein factors required to initiate translation • protein processing & degradation: • regulation of the modification or transporting of a protein • ubiquitin-tagged proteins are degraded by proteasomes
Genes Associated With Cancer • proto-oncogenes = normal cellular genes that code for proteins that stimulate normal cell growth & division • can be converted to oncogenes (cancer-causing genes) by: • translocation – proto-oncogene ends up near an active promoter or vice versa • amplification – increases # of proto-oncogenes in cell • point mutations in a promoter, enhancer, or the coding region itself • tumor-suppressor genes = encode proteins that help prevent uncontrolled cell growth • mutations that decrease the normal activity of these genes may contribute to the onset of cancer
Example: ras gene & p53 gene • ras gene is a proto-oncogene • it encodes a G protein that, in response to growth factors, triggers a cell signaling pathway that synthesizes a cell cycle stimulating protein • mutations in this gene can lead to the production of a hyperactive G protein that continuously triggers this pathway even when growth factors are absent • p53 gene is a tumor-suppressor gene • it promotes the synthesis of cell cycle inhibiting proteins • a mutation that knocks out this gene can lead to excessive cell growth & cancer
Model of Cancer Development • changes that must occur at the DNA level for a cell to become fully cancerous: • appearance of at least one oncogene • mutation or loss of several tumor-suppressor genes • in most cases mutations must knock out both alleles to block tumor suppression
Tumor Viruses • can transform cells into cancer cells through the integration of viral nucleic acid into host cell DNA • retroviruses may donate an oncogene to the cell • integrated viral DNA may disrupt a tumor-suppressor gene or convert a proto-oncogene to an oncogene • produce proteins that inactivate p53 and other tumor-suppressor proteins
Predisposition to Cancer • an individual inheriting an oncogene or a mutant allele of a tumor-suppressor gene is one step closer to accumulating the necessary mutations for cancer to develop • (ex) breast cancer genes: BRCA1 or BRCA2 • a woman who inherits one mutant BRCA1 allele has a 60% chance of developing breast cancer before the age of 50
Eukaryotic vs. Prokaryotic Genomes Eukaryotes Prokaryotes • larger genome but fewer genes in a given length of DNA • more noncoding DNA (10,000 times as much as prokaryotes ) • most of the DNA does not encode protein or RNA • genes contain introns so they are much longer than prokaryotic genes • smaller genome but more genes in a given length of DNA • less noncoding DNA • most of the DNA codes for protein, tRNA, or rRNA • genes are not interrupted by introns
Human Genome • 98.5% does not code for proteins, rRNAs, or tRNAs • 24% is gene-related regulatory sequences and introns • 44% is repetitive DNA made up of transposable elements & related sequences
Transposable Elements • two types: • transposons – move within a genome by means of a DNA intermediate • can move by “cut-and-paste” or “copy-and-paste” • retrotransposons – move within a genome by means of an RNA intermediate • always leave a copy at the original site because they are initially transcribed into an RNA intermediate • to be inserted at another site, the RNA intermediate must be converted back to DNA by reverse transcriptase
Other Types of Repetitive DNA • probably arose by mistakes that occurred during DNA replication or recombination • accounts for about 15% of the human genome • about 1/3 of this consists of large-segment duplications (10,000-300,000 base-pairs) • long stretches of DNA that have been copied from one chromosomal location to another • remaining 2/3 is simple sequence DNA – copies of tandemly repeated short sequences like GTTAC • most is located at chromosomal telomeres and centromeres indicating it has a structural role
Gene-Related DNA • consists of coding & noncoding DNA • constitutes about 25% of the human genome • about ½ of the total coding DNA consists of solitary genes • remaining ½ occurs in multigene families = collections of identical or similar genes • (ex) identical gene family = rRNA • allows for quick production of millions of ribosomes • (ex) non-identical families = α-globin and β-globin
Evolution of Genomes • extra sets of chromosomes can arise from: • errors during meiosis • polyploidy – extra sets of genes • mutations can accumulate in the extra sets of genes • unequal crossing-over • can result in one chromosome with a deletion and another with a duplication • errors during DNA replication • slippage • occurs when DNA template shifts with respect to the new complementary strand resulting in a region of the DNA not be copied or being copied twice
rearrangement of existing DNA sequences by: • exon duplication • exon shuffling • mixing & matching of different exons either within a gene or between two nonallelic genes owing to errors in meiotic recombination • transposable elements: • can promote recombination • disrupt cellular genes or control elements • carry entire genes or individual exons to new locations
Evolution of Globin Genes • all the α-globin and β-globin genes likely evolved from one common ancestral globin gene • the ancestral globin gene duplicated and diverged into α-globin and β-globin ancestral genes • the ancestral α-globin and β-globin genes later duplicated several times and their copies diverged into the current family genes • the divergences undoubtedly arose from accumulated mutations • some of the gene duplications and subsequent divergences are also suspected to have produced new genes with novel, yet related functions