Download

1 / 70
700 likes | 732 Views
Learn about the phylogenetic tree of Archaea, their energy conservation, autotrophy, and metabolism. Dive into the Phyla Euryarchaeota, halophilic Archaea, methanogens, Thermoplasmatales, and hyperthermophilic Euryarchaeota. Understand the unique characteristics and habitats of these diverse Archaea groups.

E N D
CHAPTER 13 Prokaryotic Diversity: The Archaea
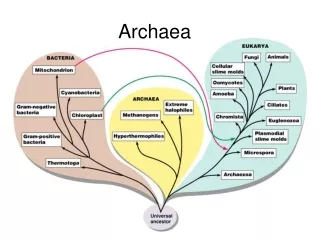
PART I Phylogeny and General MetabolismPhylogenetic Overview of the Archaea
Archaea form four major phyla, the Euryarchaeota, the Crenarchaeota, the Korarchaeota, and the Nanoarchaeota.
Energy Conservation and Autotrophy in Archaea With the exception of methanogenesis, bioenergetics and intermediary metabolism in species of Archaea are much the same as those in various species of Bacteria.
Several Archaea are chemoorganotrophic and thus use organic compounds as energy sources for growth. Chemolithotrophy is also well established in the Archaea, with H2 being a common electron donor.
The capacity for autotrophy is widespread in the Archaea and occurs by several different pathways. In methanogens, and presumably in most chemolithotrophic hyperthermophiles, CO2 is incorporated via the acetyl-CoA pathway or some modification thereof.
Extremely halophilicArchaea require large amounts of NaCl for growth. These organisms accumulate high levels ofKCl in their cytoplasm as a compatible solute.
These salts affect cell wall stability and enzyme activity. The light-mediated proton pump bacteriorhodopsin helps extreme halophiles make ATP (Figure 13.4).
Model for the mechanism of bacteriorhodopsin activity Light near 570 nm converts the protonated retinal bacteriorhodopsin from the trans form (RetT) to the cis form (RetC), along with translocation of a proton to the outer surface of the membrane, thus establishing a proton motive force. ATPase activity is driven by proton motive force. Chlorophyll pigments also synthesize ATP, a light driven process
Table 13.1 gives the ionic composition of some highly saline environments.
Table 13.2 lists the currently recognized species of extremely halophilic Archaea.
Methane-Producing Archaea: Methanogens A large number of Euryarchaeota produce methane (CH4) as an integral part of their energy metabolism. Such organisms are called methanogens. Methanogenic Archaea are strictly anaerobic prokaryotes.
Table 13.5 gives characteristics of some methanogenic Archaea.
Substrates converted to methane by various methanogenic Archaea are listed in Table 13.6. Acetotrophic substrates are those that consume acetate.
Thermoplasmatales: Thermoplasma, Ferroplasma, and Picrophilus
Thermoplasma, Ferroplasma, and Picrophilus are extremely acidophilic thermophiles that form their own phylogenetic family of Archaea inhabiting coal refuse piles and highly acidic solfataras.
Cells of Thermoplasma and Ferroplasmalack cell walls and thus resemble the mycoplasmas in this regard.
To survive the osmotic stresses of life without a cell wall and to withstand the dual environmental extremes of low pH and high temperature, Thermoplasma has evolved a unique cell membrane structure (Figure 13.11).
Structure of the tetraether lipoglycan of Thermoplasms acidophilum – monolayer of lipid rather than bilayer membrane
Hyperthermophilic Euryarchaeota: Thermococcales and Methanopyrus
Thermococcus is a spherical hyperthermophilic euryarchaeote indigenous to anoxic thermal waters in various locations throughout the world. A few euryarchaeotes thrive in thermal environments, and some are hyperthermophiles. All organisms in this group have growth temperature optima above 80°C.
Methanopyrus is a rod-shaped hyperthermophilic methanogen isolated from sediments near submarine hydrothermal vents and from the walls of "black smoker" hydrothermal vent chimneys.
Methanopyrus is unusual because it contains membrane lipids found in no other known organism. In the lipids of Archaea, the glycerol side chains contain phytanyl rather than fatty acids bonded in ether linkage to the glycerol.
In Methanopyrus, this ether-linked lipid is an unsaturated form of the otherwise saturated dibiphytanyl tetraethers found in other hyperthermophilic Archaea (Figure 13.13).
Methanopyrus produces CH4 from CO2 and H2 Unsaturated phytanyl, Geranylgeraniol produced by Methanopyrus for cell membranes
Archaeoglobus was isolated from hot marine sediments near hydrothermal vents. In its metabolism, Archaeoglobus couples the oxidation of H2, lactate, pyruvate, glucose, or complex organic compounds to the reduction of sulfate to sulfide.
Ferroglobus is related to Archaeoglobus but is not a sulfate-reducing bacterium. Instead, Ferroglobus is an iron-oxidizing chemolithotrophic autotroph, conserving energy from the oxidation of Fe2+ to Fe3+ coupled to the reduction of NO3– to NO2– plus NO (see Table 13.8).
PART III Phylum Crenarchaeota Habitats and Energy Metabolism of Crenarchaeotes
Table 13.7 summarizes the habitats of Crenarchaeota. They include very hot and very cold environments.
Most hyperthermophilic Archaea have been isolated from geothermally heated soils or waters containing elemental sulfur and sulfides. Hyperthermophilic Crenarchaeota inhabit the hottest habitats currently known to support life.
Cold-dwelling crenarchaeotes have been identified from community sampling of ribosomal RNA genes from many nonthermal environments.
Crenarchaeotes have been found in marine waters worldwide and thrive even in frigid waters and sea ice.
Hyperthermophiles from Terrestrial Volcanic Habitats: Sulfolobales and Thermoproteales
Two phylogenetically related organisms isolated from these environments include Sulfolobus and Acidianus. These genera form the heart of an order called the Sulfolobales (Table 13.9).
Key genera within the Thermoproteales are Thermoproteus, Thermofilum, and Pyrobaculum.
Hyperthermophiles from Submarine Volcanic Habitats: Desulfurococcales