Download
1 / 43
470 likes | 777 Views
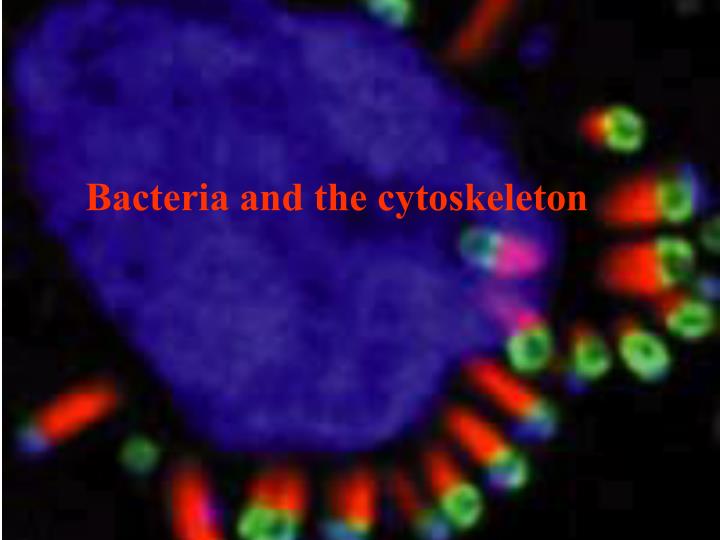
Bacteria and the cytoskeleton. The human body is a dangerous place for a bacteria to be! Antibodies Neutrophils Complement Innate response - lysozymes. Many bacteria find it much more comfortable inside the cells of its host.

E N D
The human body is a dangerous place for a bacteria to be! Antibodies Neutrophils Complement Innate response - lysozymes
Many bacteria find it much more comfortable inside the cells of its host.
Some bacteria gain entry to cells by forcing them to phagocytose them. Bacteria secrete Factors that stimulate Macrophagocytosis Once engulfed Bacteria digest the phagolysosome Now cells can grow Within the cell
Fibroblast protrusion, Louise CramerUniversity College London
GFP-actin. Stimulated Macropinocytosis
Killing phagocytosis tight compartment Stimulated phagocytosis loose compartment
Other bacteria (EPEC) stimulate the production of an elaborate adhesion (Pedestal), that prevents phagocytosis and removal by flushing
Salmonella Escherichia Yersinia Shigella Staphylococcus Legionella Listeria Many bacteria subvert normal cytoskeletal function in order to parasitize their eukaryotic host through either adhesive complexes or inducing macro-pinocytosis (Most nasty bacteria are named After people!!)
Yersinia pestis was responsible for the Great Plagues. During the 6th century AD, the plague ravaged the known world over a 50 year period causing 100 million deaths. The "black death" again devastated Europe during the 14th century over a 5 year period causing 25 million deaths (25% of the European population). The bacterium was named after Yersin who identified it as being the causative agent of an outbreak of plague in Hong Kong
Xenopsylla cheopis Dirty Rat Homo sapiens Dirty Rat Rattus norvegicus
Other Yersinia cause disease. Yersinia enterocolitica Typically, only a small number of human cases of Yersiniosis are recognized. Symptoms are like that of appendicitis and out breaks are often detected by a sudden increase in appendectomies in a particular region. The Center for Disease Control & Prevention estimates that about 17,000 cases occur each year in the United States.
Bacteria inject toxins into cells to subvert their activities The hypodermic syringes that they use are modified flagella
Type III Type II Type I Outer membrane Periplasm Inner membrane A A A The three main types of bacterial secretion Type III is most often associated with pathogenic bacteria
The most common pathogenic E.coli Abbrev. Full name Common name and features inocolum Source ETEC Enterotoxin E.coli Montizuma’s revenge, traveller’s tummy 108 Faecal usually comparatively mild, (Diacalm grade) contamination EIEC Enteroinvasive Invades, Shigella pathogenicity island high Food & waterborne EPEC Enteropathogenic Pedestal formation, infant diarrhoea 108 - 10 Nosocomial community EHEC Enterohaemorrhagic (O157)“Hamburger disease” Shiga toxin 3 Cattle faeces, meat
Interactions of the common pathogenic E.coli with epithelial cells
Interaction of EPEC with epithelium first through EspA filaments (a), then through intimin (b). Knutton et al, 1998. Nucleolin is a third binding site.
Scanning E.M. of EPEC and epithelium. EspA filaments appear to insert into cell (arrows in A), possibly to deliver EspB. EspA may be part of the Type III secretion pathway, it is needed for EspB delivery. Note the pedestals are all of equal length.
Some pathogenic E.coli (EPEC, EHEC) put down their own “Welcome mat” Tir (translocated intimin receptor) is injected into host by Type III secretion Tir binds to host a-actinin, talin and vinculin all components of the focal contact. Nucleolin is a bacterial binding site for EPEC.
Cell death & loss Lumen of gut Movement of fresh cells up from crypt Cell division By targeting nucleolin E.coli are able to attach to the cells that will exist for the longest time. E.coli bind dividing cells in the crypt and stay attached as the are conveyed to the tips.
Microfilaments are present in ‘stress fibres’that are attached to‘Focal adhesions’. They are also present as a gel under the plasma-membrane esp. at the leading edge
The pedestal has features in common with both the focal contact and microvilli Arp2/3 WASP a -actinin Vinculin Villin Ezrin Pedestal base Myosin II tropomyosin
O II + Actin NH-C-CH I H-S-H C- 2 2 Actin NH-C-CH -S-H C- 2 2 Pyrene-actin method to measure polymerisation kinetics Pyrene Excited Light emitted at at 366nm 384nm measured Pyrene-actin in quartz cuvette
Wiskott-Aldrich Syndrome Protein (WASP) A c i d i c P r o l i n e - r i c h W H 2 C I n a c t i v e N B a s i c C R I B A c i d i c P r o l i n e - r i c h W H 2 C P P A r p c o m p l e x A c t i n A c t i n A c t i v e C R I B B G a s i c N
The Arp2/3 complex An actin-binding group of proteins pivotally involved in the regulation of actin polymerisation. Z e r o l e n g t h c r o s s l i n k A r p 3 A r p 2 N o n - z e r o l e n g t h c r o s s l i n k Y e a s t t w o h y b r i d s c r e e n p 1 8 p 1 9 p 3 5 p 4 0 M i c r o f i l a m e n t b i n d i n g p 1 4
Analysis of the WASP domains required for Pedestal formation WASP-WT WASP-DC WASP-DGBD Kalman et al, 1999 Nature Cell Biol. 1; 389-391.
Pedestal formation and localization of Arp2/3 complex components. Kalman et al, 1999 Nature Cell Biol.1; 389-391.
Edwina Currie Eggs Salmonella Salmonella
Pathogenic Salmonella disrupt normal cell function Salmonella Type III secretion Rac GDP Cdc42 GDP SopE SopB SipC SipB SptP SipA RacGTP Cdc42 GTP Plastin Active WASP Actin polymerizing and bundling Arp2/3 complex Inactive WASP
Hints of Plastin’s involvement in signalling & Bacterial invasion. 1). BPB inhibition of plastin inhibits IP3 dependent Ca2+ increase in PMNs. 2). Plastin is itself regulated by Ca2+. 3). Phosphorylation of plastin at Ser5 by PKA results in integrin activation in PMNs stimulated by Fc receptor ligation
Pathogenic Salmonella disrupt normal cell function Salmonella Type III secretion Rac GDP Cdc42 GDP SopE SopB SipC SipB SptP SipA RacGTP Cdc42 GTP Plastin Active WASP Actin polymerizing and bundling Arp2/3 complex Inactive WASP
The Salmonella cycle of infection SPI1 SPI2 Initial contact A fresh actin wave of actin polymerization results in the vacuole being covered in actin. Actin polymerization and phagocytosis Injection By type III secretion Phagosome stimulates new protein secretion Through a second type III machine Lysosomes can’t fuse Nucleus Some time later an actin ADP-ribosylating enzyme disassembles the structure for unknown reasons (cell lysis & spread?).
Gel filration of SipC SipC and actin SipC Actin SipC and actin (Higher power) SipC and actin Hayward, R.D. & Koronakis, V. 1999 EMBO J.18, 4926-4934.
Actin + SipC-C Actin + SipC-N Gel filtration Actin + SipC-C
Co-sedimentation of SipC N-terminus with actin bundles. Sedimentation of actin bundles from a mixture of SipC-N and F-actin (both 5 µM), demonstrating formation of an actin-SipC-N complex. Supernatants (S) and pellets (P) after centrifugation
Actin+SipC-C Actin Actin+SipC-C+ Cyto Actin + Cyto SipC C-terminal domain SipC-C inhibition by Cytochalasin D E.M. of actin with SipC-C and SipC-N Bundle
Induction of cytoskeletal rearrangements in vivo by SipC and SipC-C microinjection. Cultured HeLa cells fixed 30 min after microinjection with purified SipC (upper panels) or SipC-C (lower panels) (3 µM). Cells (DIC; A and D) were stained with polyclonal antibody to SipC and FITC-conjugated anti-rabbit IgG [SipC (B), SipC-C (E)] and with Texas Red-conjugated phalloidin to visualize F-actin [SipC (C), SipC-C (F)]. Injected cells are indicated by arrows.
Co-injection of SipC-N with GST-GFP. Cultured HeLa cells (DIC; A and D) fixed 20 min after microinjection with GST-GFP alone (upper panels) or mixed with SipC-N (lower panels) (3 µM). GST-GFP was visualized directly [GST-GFP alone (B); + SipC-N (E)] and F-actin stained with Texas Red-conjugated phalloidin [GST-GFP alone (C); + SipC-N (F)]. Injected cells are indicated by arrows (N = nuclear injection).
The End Be Free