Download

1 / 34
600 likes | 1.5k Views
The Cytoskeleton. The cytoskeleton is an aggregate structure formed of 3 kinds of cytoplasmic filamentous proteins: microfilaments (actins), intermediate filaments and microtubules (tubulins). I. Actin-based cytoskeleton. Positioning of proteins and organelles Cell shape – all cells

E N D
The Cytoskeleton The cytoskeleton is an aggregate structure formed of 3 kinds of cytoplasmic filamentous proteins: microfilaments (actins), intermediate filaments and microtubules (tubulins)
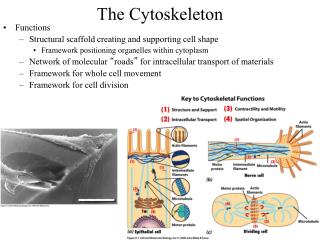
I. Actin-based cytoskeleton • Positioning of proteins and organelles • Cell shape – all cells • Shape changes • Ameboid locomotion • Neuronal growth cones • Muscle contraction
Microfilaments: visualized in a kidney cell with fluorescently-labeled phalloidin (a poisonous mushroom toxin that stabilizes filaments and inhibits processes that depend on actin filament turnover)
Actin filaments have polarity, a fact that is relevant for filament growth and interaction with other cellular elements + or ‘barbed’ ends - or ‘pointed’ ends
Assembly of microfilaments Monomers of G (globular) actin polymerize to form long filaments, the F (filamentous) actin. The addition of actin monomers to the ends of the filament occurs at different rates: addition to the + end, also called the barbed end occurs 5-10x faster than to the minus or pointed end. Polymerization requires hydrolysis of an ATP.
Reversible polymerization of actin: Treadmilling of actin monomers(In some cells, remodeling of actin filaments is responsible for half of the ATP hydrolysis in the cell)
Growth of a microfilament requires an initial nucleation by an actin trimer, and can stop if the rate constant for adding monomers at the + end and one for removing them at the – end approach the same value – at that point, treadmilling occurs.
Some intracellular pathogens such as the bacteria Listeria and Shigella and the vaccinia virus usurp the host cell’s mechanism of assembling actin networks and propel themselves through the cytoplasm with actin “tails”.
Vaccina viruses can use the cytoskeleton of infected cells to spread within the body, and at the same time prevent already-infected cells from becoming super-infected. Once a cell is infected, the virions produce 2 proteins that act at the cell surface to induce formation of actin tails. Virions are then propelled from the infected cell on the tip of these actin tails and can land on and infect nearby cells that are not infected. If a virion falls back on the same cell or another infected cell, it gets relaunched. An infected cell could be juggling hundreds of virions at any instant. (Science 327, 787 (2010))
Association of actin filaments with the membrane • A network of actin filaments and other cytoskeletal proteins underlies the plasma membrane and determines cell shape: example, rbc: • Actin bundles also attach to the plasma membrane to anchor cell-cell and cell-substratum contacts, as shown here:
Microvilli in an intestinal epithelial cell • Each of the ~ 1000 microvilli contain several dozen microfilaments with their + ends facing outward; they are tightly packed together by actin-bundling proteins and are connected to the inner surface of the plasma membrane by lateral crosslinks composed of calmodulin and myosin 1. They project into the dense actin network within the cell.
Structure of microvilli • Microvilli are relatively stable projections of the plasma membrane that are supported by microfilaments. Microvilli increase the surface area of cells such as those lining the intestine.
The actin cytoskeleton is 1) linked to the plasma membrane and2) able to change to promote cell movement • Cytochalasins (toxins that are synthesized by fungi) bind to the + ends and block elongation. This blocks pseudopodial extension and cleavage furrow formation after mitosis. • In this picture of a resting, attached fibroblast, the long filaments are revealed by phalloidin staining, but when a fibroblast begins to move, the filaments break down into short filament fragments and G actin and filaments are reassembled in the cell’s extensions.
The mechanical properties of cytoplasm are regulated by crosslinking of actin: slow pressure allows adaptation in the connections whereas rapid deformations are resisted
Actin filament turnover and monomer recycling is regulated by accessory proteins
Actin exists in cells as bundles and networks: actin-binding proteins crosslink the filaments
Extrinsic signals can regulate the actin cytoskeleton • Phosphoinositol bis phosphate (PIP2) binds profilin, preventing if from interacting with actin. • Phospholipase C hydrolyzes PIP2 to IP3, releasing profilin, which promotes filament growth • Gelsolin and cofilin are actin-severing proteins – they are inhibited by PIP2 - when they are active, they stimulate formation of new filaments by increasing the number of + ends available
Several g-protein coupled pathways control cell shape and behavior • The Rho protein family includes the small monomeric G proteins Cdc 42, Rac and Rho • The Cdc42 pathway activates rapid actin polymerization and bundling, causing cells to put out filopodia • The Rac pathway uncaps + ends of filaments, stimulating filament elongation, causing cells to form lamellipodia and membrane ruffles • The Rho pathway stimulates filament bundling, causing cells to form stress fibers that interact with focal contact points with the extracellular matrix
Fibroblast cell shape changes resulting from single g-protein signals Dynamic interaction of these signals is necessary for complex cellular behavior, as seen in the next slide of cell crawling.
Dynamics of actin in a crawling cell: note the polarization of the microfilaments
II. Intermediate filaments • Formed by a large, heterogeneous group of proteins At least 4 major filament types • Keratin - epithelial cells • Neurofilaments – neurons • Vimentin-containing filaments – fibroblasts, glial cells, muscle cells • Nuclear lamina – all nucleated cells • No energy is required for filament assembly • Filaments are not polarized
Intermediate filaments in kidney cell (left, green) and cultured epithelial cell (right, orange; desmosomes are green)
Genetic diseases of skin and nervous system • Mutant keratin genes are responsible for some human skin diseases – some lead to susceptibility to skin damage and death due to the trauma of birth, others impact the organization and stability of skin. • Abnormalities of neurofilaments underlie the loss of motor control in Lou Gehrig’s disease (ALS) and the motor neuron disease that afflicts the physicist Stephen Hawking.
III. Microtubule-based cytoskeleton • Microtubules are polymers of tubulin • Microtubules are dynamically unstable – they can be assembled and disassembled rapidly • Tubules are organized by the centrosome complex • Functions: • Vesicle and organelle transport (ex. Fast axoplasmic transport) • Cilia and flagella • Mitotic spindles
III. Microtubules: α,β,γ tubulin • Microtubules are stiff, cylindrical polymers of α and β tubulin. • The polymers project from the centrosome, where the γ tubulin is present that is necessary for the “nucleation” of the polymers. • There is a + end projecting into the cytoplasm and a – end that often remains anchored in the centrosome. (cilia and flagella are a special case, as we will see).
Regulation of growth and dissolution by subunit supply: high and low concentrations of GTP-tubulin
Compared with the microtubules of a normal fibroblast (A), the cell in (B) is regrowing microtubules from the centrosome after treatment with colcemid blocked assembly of microtubules
Chemicals that affect microtubules: drugs that inhibit mitosis by having opposite effects • Colchicine (Colcemid) is an alkaloid from the autumn crocus that binds to tubulin monomers, inhibiting their assembly. (In the previous slide, the disappearance of the microtubules resulted from 60 min. exposure to colcemid, which shows that continuous disassembly and reassembly underlies the normal presence of the filaments.) Nocodazole, (a synthetic benzimidazole) is an experimental drug that inhibits microtubule assembly and is more rapidly reversible than colchicine. • Vinblastine and vincristine are alkaloids from the periwinkle plant, Vinca, that are used in cancer chemotherapy because they cause tubulin to aggregate. • Taxol (from the Pacific yew tree) binds to tubules and stabilizes them, making it an anticancer drug (for breast and ovarian cancers) because it inhibits cell division.
Like microfilaments, microtubules in the cytoplasm exhibit dynamic instability, including treadmilling and motility that results from the pattern of growth, such as growing towards and then moving chromosomes in mitosis.
Putting them all together: microfilaments, intermediate filaments and microtubules in kidney cells
Microtubules, intermediate filaments and the actin filaments (gray) are all linked by a protein called plectin (orange, with gold particles on the strands)