Download

1 / 20
200 likes | 330 Views
SBS922 Membrane Biochemistry. Mitochondria and chloroplasts. John F. Allen School of Biological and Chemical Sciences, Queen Mary, University of London. 1. http://jfa.bio.qmul.ac.uk/lectures/. School of Biological and Chemical Sciences Seminars 2006-07

E N D
SBS922 Membrane Biochemistry Mitochondria and chloroplasts John F. Allen School of Biological and Chemical Sciences, Queen Mary, University of London 1
School of Biological and Chemical Sciences Seminars 2006-07 WEDNESDAYS AT 12 NOON IN LECTURE THEATRE G23, FOGG BUILDING, SCHOOL OF BIOLOGICAL AND CHEMICAL SCIENCES
From these sorts of measurements observed characteristics fall into 5 main categories 1] Energy coupling sites 2] Respiratory control 3] Uncoupling agents 4] Phosphorylation inhibitors 5] Reverse electron transfer You will observe 1-4 in Practical 4.
2] Respiratory Control In 1956 Chance and Williams observed that “as long as substrate (reductant, source electrons, electron donor), oxygen (oxidant, electron acceptor) and phosphate not limiting, rate of electron transfer effectively controlled by availability of ADP” i.e. whether ATP synthesis taking place or not. This phenomenon called respiratory control a) in absence of ADP, rate of electron transfer is low, reflects slow rate at which energy for ATP synthesis (provided by electron transfer) is dissipated in absence of ATP synthesis. Called “controlled state” or state IV b) if then add ADP the rate of electron transfer will increase dramatically until rate (state III) controlled by rate at which energy diissipated (actually limiting factor is activity of ADP/ATP translocase importing ADP and exporting ATP) c) rate then remains fast until almost all ADP phosphorylated to ATP. At this point rate declines to state IV again
So electron tranfer rate and ATP synthesis tightly coupled to each other in intact mitochondria i.e. rate of electron transfer controlled by rate of ATP synthesis so that under normal conditions electron transfer is not releasing redox energy at such a rate that it is wasted before ATP synthesis can take place. Can calculate a respiratory control ratio State III/ State IV. This is a sensitive guide to “tightness” of coupling between e.t. and ATP synthesis (value rarely exceeds 7). If damage coupling membrane will decrease, so any theory will have to explain why need intact membrane for efficient coupling. Can use respiratory control to determine ADP/O quotient and you will do so in practical 3. Energy released by electron transfer can drive other energy-dependent functions such as active transport, if add Ca2+ to mitochondria in State IV can induce increase in electron transfer as energy used for active transport.
3] Uncoupling agents Large number of compounds, when added to mitochondria a) abolish ATP synthesis b) abolish other energy-dependent membrane-linked functions such as active transport c) at the same time cause a permanent stimulation in rate of electron transfer, to a rate greater than state III, whether ADP and Pi present or not These compounds called uncoupling agents, and mitochondria said to be uncoupled. Uncouplers seem to affect each of individual coupling sites to same extent. Conclusion is that electron transfer coupled to ATP synthesis via an energised state, which is non-phosphorylated, and common to all coupling sites. Uncouplers dissipate this energised state as heat, and no ATP synthesis takes place. Electron transfer at maximal rate because no longer controlled by back-pressure from energised state.
4] Phosphorylation inhibitors These actually inhibit activity of ATP synthase so preventing use of energised state for ATP synthesis. They have the following effects a) cause ADP-induced sate III rate to decrease back to state IV and further additions of ADP have no effect b) have no effect on uncoupled rate of electron transfer c) if mitochondria already inhibited to state IV by phosphorylation inhibitor, added uncoupler will stimulate rate of electron transfer. This evidence strongly suggests that uncouplers act at a point between electron transfer and ATP synthesis i.e. energised state.
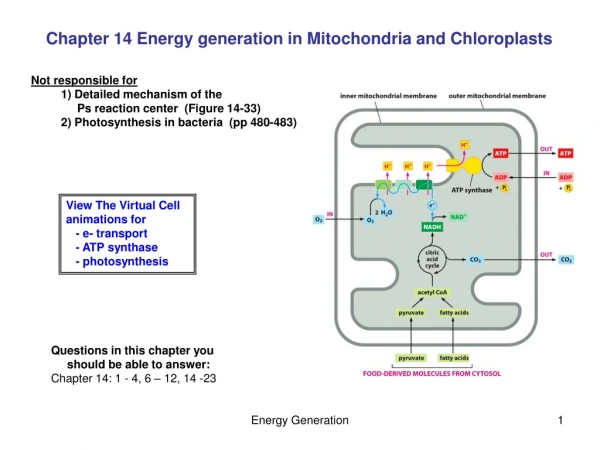
Mitchell’s chemiosmotic hypothesis Below shows old fashioned (late 1950's/early 1960's) view of mitochondrial electron transfer as a series of redox (oxidation-reduction) carriers in the inner mitochondrial membrane. This view neglects entirely the significance of the proteins that nearly all these redox components are bound to in the membrane.
A MORE MODERN VIEW OF MITOCHONDRIAL ELECTRON TRANSFER AND ATP SYNTHESIS ACKNOWLEDGES THE IMPORTANCE OF THE RESPIRATORY CHAIN COMPLEXES. COMPLEX O )ENZYME NAME / NICOTINAMIDE NUCLEOTIDE TRANSHYDROGENASE )LOCATION COUPLING SITE 0
COMPLEX I )ENZYME NAME/ NADH-UBIQUINONE(UQ) OXIDOREDUCTASE )LOCATION OF COUPLING SITE 1 COMPLEX II )ENZYME NAME/ SUCCINATE-UQ OXIDOREDUCTASE )NO COUPLING SITE
COMPLEX III )ENZYME NAME/ UBIQUINOL(UQH2)-FERRICYTOCHROME C3+ OXIDOREDUCTASE )LOCATION COUPLING SITE 2 COMPLEX IV )ENZYME NAME/ FERROCYTOCHROME C2+-OXYGEN OXIDOREDUCTASE )LOCATION COUPLING SITE 3
COMPLEX V )ENZYME NAME/ ATP SYNTHASE The handouts and the list above also show the location of coupling sites (sites where electron transfer through the chain at that point is linked to ATP synthesis) in the mitochondrial electron transfer chain in relation to the points at which substrates donate electrons to the chain.