Download
1 / 50
500 likes | 730 Views
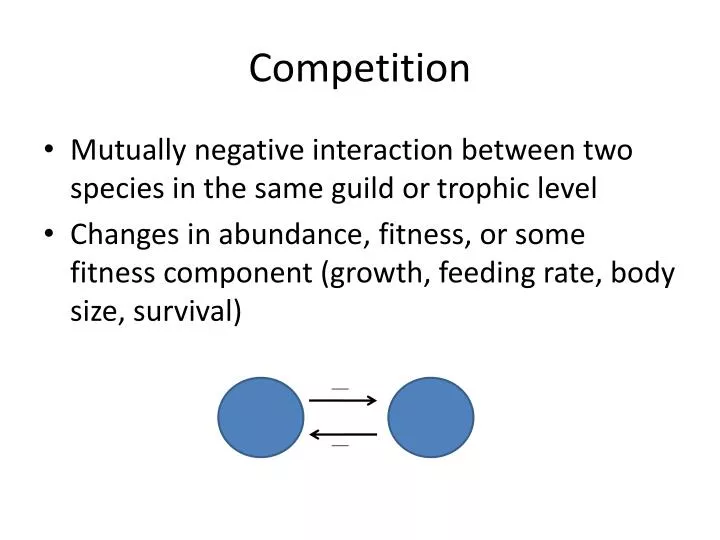
Competition. Mutually negative interaction between two species in the same guild or trophic level Changes in abundance, fitness, or some fitness component (growth, feeding rate, body size, survival). Topics for today. Mechanisms and models of competition Evidence for competition

E N D
Competition • Mutually negative interaction between two species in the same guild or trophic level • Changes in abundance, fitness, or some fitness component (growth, feeding rate, body size, survival)
Topics for today • Mechanisms and models of competition • Evidence for competition • Experiments (lab, field) • Observational (competitive exclusion, character assortment or displacement) • Latest advances in the study of competition • Genetic diversity/distance and competition • Functional trait complementarity and competition • Habitat filtering vs. competitive exclusion using phylogenetic methods
Schoener (1983): Mechanisms • Consumption • Pre-emption • Overgrowth • Chemical interactions • Territoriality • Encounter competition • Can we think of other sorts?
Lotka-Volterra models of competition • Based on estimates of logistic population growth, and how this differs in monoculture vs. mixtures
Values of population sizes of two species, N1 and N2, that result in positive, negative, or zero population growth for interacting species. The zero growth isoclines are shown as a solid line for species 1 and a dashed line for species 2. [from Morin 1999] K is carrying capacity; dN/dt is population growth rate; a is competition coefficient with a12 being effect of sp. 2 on sp.1
Species 2 Species 1 Figures from Gotelli, “A Primer of Ecology”
Competitive Exclusion ...of species 2 by species 1 ...of species 1 by species 2 Figs from Gotelli
Equilibrium: stable coexistence vs. unstable competitive exclusion “winner” depends on priority effects coexistence Figs from Gotelli
R2 R1 Tilman’s mechanistic model R2B R2A R1A R1B
Tilman’s model • If ZNGI’s overlap, we add consumption vectors (C) to illustrate how each species uses resources • If each species consumes more of the resource that limits itself, get coexistence • If each species uses more of the resource that limits the other species, outcome is unstable
R2 R1 Tilman’s model
R2 R1 Tilman’s model
Competitive ability can be measured by species traits in monoculture • R*: the amount of resource left when a population of a single species reaches equilibrium density • Species with lowest R* should competitively exclude all others
Evidence for competitive ability:Tilman’smeasure of R* Poor competitors remove less N Good competitors remove more N Roots are the foraging organ: mass correlated with N assimilation Tilman and Wedin 1991
Tilman’s field expt. Wedin and Tilman 1993.
Connell’s barnacle experiment Chthamalus stellatus Nucella = Thais lapillus Balanus balanoides
Hairston’s salamanders Hairston 1980
Anoles in the Lesser Antilles Similar body size and perch height Anolisgingivinus Little overlap in size or perch height Anoliswattsi Anolisbimaculatus
Treatments with “W”: competing with A. wattsi Similar niche Different niche Pacala and Roughgarden 1982 High body size and perch height overlap results in competition
Patterns from field experiments • Are there traits that predict who ‘wins’? • Are there traits or patterns that predict where competition is more intense?
Observational evidence for competitive exclusion: MacArthur’s Warblers
Galapagos finch bill sizes differ more in sympatry than allopatry
But differences in bill size between co-occurring pairs no different than expected by chance?? Strong et al 1979
Yet the Grants showed that evolution did indeed occur Large-beaked G. fortis (A) and G. magnirostris (B) can crack or tear the woody tissues of T. cistoidesmericarps (D), whereas small-beaked G. fortis (C) cannot.
Drought selects for larger beaks in fortis (only large seeds available) Drought causes competition: selects for divergent (small) beaks in fortis magnirostris introduced: has really large beak Grant and Grant 2006
Desert cats DO show character displacement when tested against null models Canine size for each species/sex in two locations Dayan et al 1990
And so do bat-pollinated Burmeistera Muchhala and Potts 2007
Current areas of inquiry • Is competition stronger for closely related species? And, can we infer whether competitive exclusion has occurred using phylogenetic methods? • Is competition stronger for species in the same functional group?
Darwin 1859 “As species of the same genus have usually, though by no means invariably, some similarity in habits and constitution, and always in structure, the struggle will generally be more severe between species of the same genus, when they come into competition with each other, than between species of distinct genera.”
If habitats select for particular traits, and related species share traits, expect phylogenetic clustering (“habitat filtering”) If competition or other density-dependent factors are stronger between relatives than between distant relatives, expect phylogeneticoverdispersion
Two different estimates of relatedness Webb 2000: evidence for habitat filtering in tropical trees NRI: species more related than expected (clustering); NTI: not different from random (do NOT see overdispersion)
Cavender-Bares et al 2004: evidence for competitive exclusion in oaks
Using experimental estimates of competition, closely related species do not compete more intensely Cahill et al 2008 These are correlations between competitive effect and phylogenetic distance. What do you expect this correlation to be if closely related things compete more intensely?
In general.... • Productivity is greater in plots with higher species richness • Is this because competition is lower in diverse plots (more functional groups present)?
Two hypotheses • Niche Complementarity: different functional groups use resources differently (in “complementary” ways), so greater efficiency • Selection Effect: with higher diversity, greater chance of “selecting” a competitive dominant in a plot (ie species that grow large over time) Fargione et al 2006
Net effect: Difference between total biomass in a plot and average biomass of monocultures increases over time Complementarity: when positive (as here) means species have higher than expected yield in mixture (attributed to N-fixers and C4 species presence in high-diversity mixtures) Selection: if positive would mean that species with high monoculture biomass are competitive dominants, and when present (by chance) create more total biomass in diverse plots Fargione et al 2006
Is it really about functional group diversity, or another diversity metric? Cadotte et al 2009
Main points • Models as a way to think about what we can measure in the field • Experiments can show patterns of functional traits that are important • Phylogenetic inference can provide new insight for experimentally intractable systems, and new interpretation of data from others
Reading for next week • Connell 1961
Booth, R. E., and J. P. Grime. 2003. Effects of genetic impoverishment on plant community diversity. Journal of Ecology 91:721–730. • Cadotte, M. W., J. Cavender-Bares, D. Tilman and T.H. Oakley. 2009. Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity. PLoS ONE 4(5): e5695. • Cavender‐Bares, J., D. D. Ackerly, D. A. Baum, F. A. Bazzaz. 2004. PhylogeneticOverdispersion in Floridian Oak Communities. Am Nat 163:823-843 • Connell, J. H. 1961. The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalusstellatus. Ecology 42: 710-723. • Connell, J. H. 1980. Diversity and the Coevolution of Competitors, or the Ghost of Competition Past. Oikos 35:131-138. • Dayan, T. , D. Simberloff. 2005Ecological and community-wide character displacement: the next generation. Ecology Letters 8: 875-894. • Dayan, T., D. Simberloff, E. Tchernov, Y Yom-Tov. 1990. Feline canines: community-wide character displacement among the small cats of Israel. Am. Nat. 136: 39-60. • Fargione, J.; Tilman, D. 2006. Predicting relative yield and abundance in competition with plant species traits. Functional Ecology 20:533-450. • GauseG. F. 1934. The struggle for existence. Williams & Wilkins. • Grant, P.R. and B. R. Grant. 2006. Evolution of character displacement in Darwin’s finches. Science 313: 224-226. • Hairston N. G. 1980a The experimental test of an analysis of field distributions: competition in terrestrial salamanders. Ecology 61: 817-826. • JF Cahill, SW Kembel, EG Lamb, and PA Keddy. 2008. Does phylogenetic relatedness influence the strength of competition among vascular plants? Perspectives in Plant Ecology, Evolution, and Systematics 10:41-50. • Macarthur R.H. 1958. Population ecology of some warblers of northeastern coniferous forests. Ecology 39: 599-619. • Muchhala, N. And M. D. Potts. 2007. Character displacement among bat-pollinated flowers of the genus Burmeistera: analysis of mechanism, process and pattern. Proc Roy Soc B 274: 2731-2737 • SchoenerT. W. 1983. Field experiments on interspecific competition. Am Nat 122: 240-285. • Strong, D. R., L. A. Szyska, D. Simberloff. 1979. Tests of community-wide character displacement against null hypotheses. Evolution 33: 897-913. • Tilman, D. and D. Wedin. 1991. Plant traits and resource reduction for five grasses growing on a nitrogen gradient Ecology 72: 72:685-700 • Webb CO. 2000. Exploring the phylogenetic structure of ecological communities: an example for rain forest trees. Am. Nat. 156:145– 55 • Wedin D. And D. Tilman. 1993. Competition Among Grasses Along a Nitrogen Gradient: Initial Conditions and Mechanisms of Competition. Ecological Monographs 6:3 199-229