Download
1 / 26
520 likes | 2.2k Views
Cell-Mediated Immunity. An adaptive immune response mediated by specific cells of the immune system Primarily T lymphocytes ( T cells ), but also macrophages and NK cells .

E N D
Cell-Mediated Immunity • An adaptive immune response mediated by specific cells of the immune system • Primarily T lymphocytes (T cells), but also macrophages and NK cells. • Formally defined as immunity that can be transferred from one organism to another by lymphoid cells, but not by serum antibody.
T cells • Main coordinators and effectors of cellular immunity • Defined by their development in the thymus and the presence of a T-cell receptor (TCR) complex
T cells (continued) • Two main types: 1. CD4+: Stimulate other immune cells. 2. CD8+Cytotoxic T cells: Kill intracellularly-infected cells. • Two major types of CD4+ T cells: 1. TH1: Inflammatory T cells -- Stimulate macrophages and promote inflammatory responses. 2. TH2: Helper T cells -- Stimulate B-cells to produce antibodies. (A third type, TH3, has recently been shown to promote IgA production.)
T cells develop in the thymusand undergo positive and negative selection • Positive selection: T cells which can react to self MHC (major histocompatability complex) carrying peptides are allowed to live. Those that cannot undergo apoptosis (suicide). • Negative selection: T cells that react strongly to self-antigens on MHC are eliminated. • Only those T cells that can react to MHC, but do not bind strongly to self-antigens emerge as mature T cells from the thymus. • Only about 2% of immature T cells make it through positive and negative selection.
T cell development T cell development in the thymus: Immature double-negative T cells (CD8-, CD4-) Positive selection/ negative selection Immature double-positive T cells (CD8+, CD4+) Cortex Medulla CD8+ T cells CD4+ T cells Mature T cells
Epitope-binding site chain chain Variable region Constant region Transmembrane region The T cell Receptor • Similar in structure to Immunoglobulins (similar to a single Fab fragment. • Composed of two glycoprotein chains (/ or /). Most mature T cells have TCRs composed of an chain and a chain (they are called / T cells). • Each chain has a constant region and a variable region, similar to an antibody light chain. • A TCR recognizes a small (8-13 aa) peptide epitope displayed on MHC
TCR compared to Immunoglobulins Similarities • Both have specific Antigen-binding region created by the variable regions of two polypeptide chains. • Both display great potential for diversity via genetic recombination at the genome level Differences • A TCR is monovalent (has one binding site). An Ig is bivalent (has two binding sites). • The TCR has no secreted form. It is always membrane-bound. • The TCR does not recognize free antigen. Antigen must be presented to a T cell on an MHC molecule (next week). • There is no class switching for the TCR. Once made, the TCR does not change. Epitope-binding site chain chain Variable region Constant region Transmembrane region Immunoglobulin T cell Receptor
TCR: Antigen recognition CD3 CD3 Cell signaling The T cell Receptor, cont. • The TCR only recognizes specific peptide/MHC complexes expressed on the surfaces of cells • A TCR complex is composed of one heterodimeric TCR (ususally /), plus a 5-polypeptide CD3 complex which is involved in cell signalling for T cell activation. • Each TCR is produced through genetic recombination and recognizes one small peptide epitope (about 8-13 amino acids). • One T cell expresses only one specific type of TCR. CD3 is the activation complex for the TCR Binding of antigen/MHC to the TCR stimulates CD3. CD3 then sends an activation signal to the inside of the T cell.
This chart is not intended to be memorized Responses to infection -- T cell component Recognition by pre-formed, non- specific effectors Removal of infectious agent Innate immunity (0-4 hours) Infection Recognition and activation of effector cells Early induced response (4-96 hours) Removal of infectious agent Recruitment of effector cells Infection Clonal expansion and differentiation to effector cells Late adaptive response >96 hours) Transport of antigen to lymphoid organs Recognition by naïve B and T cells Removal of infectious agent Infection Recognition by pre-formed, Ab and T cells Protective immunity Removal of infectious agent Infection Rapid expansion and differentiation to effector cells Recognition by memory B cells and T cells Immunological memory Removal of infectious agent Infection The adaptive immune response involving antigen-specific T cells and B cells is only one part of the immune response and is required to protect against pathogens. A pathogen is by definition an organism that can cause disease. In other words, a pathogen is an organism that can bypass innate immunity and requires an adaptive immune response for clearance.
Generation of an adaptive immune response • During an adaptive immune response,T cells which recognize specific antigen(s) are selected for differentiation into armed effector cells which undergo clonal expansion to produce a battery of antigen- specific cells. • Clonal expansion refers to the process by which antigen-specific T cells or B cells are stimulated to reproduce clones of themselves to increase the system’s repertoire of antigen-specific effectors.
Generation of an adaptive immune response • Activation of antigen-specific T cells (the initiation of the adaptive response) occurs in the secondary lymph tissues (lymph nodes and spleen). • This activation depends upon antigen presentation by a professional antigen presenting cell (APC) along with simultaneous co-stimulation. (eg., B7 on the APC, CD28 on the T cell).
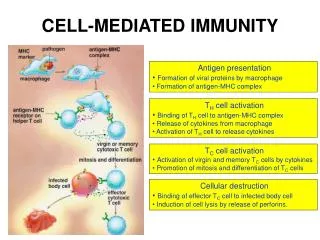
Initiation of the adaptive immune response • The first step is the draining of antigen into the lymph node(s). • In the lymph node(s) (or spleen), antigens are trapped by professional APCs which display them to T cells.
The antigen presenting cells, continued Note: this B cell is not a plasma cell -- a plasma cell is shown above. Plasma cells do not present antigen. They simply pump out antibody for a few days then die. Dendritic Cell Macrophage B cell
T cells continuously circulate via the blood and lymph through different lymph nodes until they either find presented antigen or eventually die • When a T cell encounters an APC displaying antigen to which it can bind, it stops migrating and binds strongly to the APC. • Within about 2 days (48 hours), most antigen-specific T cells have been trapped by antigen and within about 4 to5 days armed effector T cells are migrating out of the lymph node.
TH0 IL-2 IL-4 IFN- TH1 TH2 IL-2 IFN- F IL-4 IL-6 IL-10 Review -- Cytokines produced early in response to infection influence the future functions of activated CD4+ cells • Cytokines produced by TH1 cells inhibit TH2 cells • Cytokines produced by TH2 cells inhibit TH1 cells • An immune response is often dominated by a cell-mediated response or an antibody response. • Some pathogens have evolved strategies to shift the immune response toward the less effective type for that pathogen.
Functions of the different T cell types • CD8+ cells: Kill virally infected cells • CD4+ cells: • TH1: Activate macrophages to aggressively ingest antigen and to kill ingested microbes. • TH2: Stimulate B cells to differentiate into antibody-producing plasma cells. B cells will only undergo isotype switching after receiving T cell help. The Ig class that a B cell switches to is specified by the types and balance of cytokines secreted by the helper T cell.
Immunological memory • When B cells are activated to reproduce, some differentiate into plasma cells and some become long-term memory cells. • An adaptive immune response also produces T cell memory, but the nature of memory T cells is unknown. Two possibilities exist. Memory T cells probably originate from either: • 1. A long-lived subset of effector T cells that differentiates into memory T cells -- like memory B cells. • 2. The continuous low-level activation of naïve T cells by specific antigen that is retained in the lymph nodes after an infection. This mechanism would suggest that APCs in the lymph node hold on to antigen on a long-term basis after an infection and continuously stimulate T cells at a low level so there is always a small effector population ready to go.
MHC classes I and II Functions: • class I MHC: • Displays peptides derived from antigen originating inside the cell (endogenous antigen). • Important in cytotoxic responses (eg, CD8+-killing of virus-infected cells). • Class II MHC: • Displaysantigen derived from ingested antigens (exogenous antigen). • Important in humoral (antibody) responses as well in fighting as some intracellular parasites (eg. Mycobacterium tuberculosis and M. leprae) • Locations: • Class I MHC found on all nucleated cells (all cells need to be prepared to be killed in case of a viral take-over or tumorigenic transformation). • Class II MHC found only on antigen presenting cells (cells that present antigen to CD4+ T cells --> Macrophages, activated B-cells, dendritic cells.
Antigen Presentation to T cells: MHC • Antigens are presented to T cells as short peptide fragments bound to Major Histocompatibility (MHC) molecules. • Two types of MHC in humans and mice: • MHC I: presents an 8-10 amino acid peptide to CD8+ T cells. • MHC II: presents a longer peptide (13 aa or more) to CD4+ T cells.
Peptide binding cleft Peptide binding cleft 2 1 1 1 3 2-microglobin 2 2 Class I MHC Class II MHC MHC structure • MHC classes I and II have an almost identical 3-D structure. • Both classes of MHC are polygenic (each cell has many MHC genes) and polymorphic (there are many alleles for each locus), but the MHC genes do not undergo recombination. Note: Human MHC are called HLA (human leukocyte antigen).
MHC / T cell interactions Class II MHC Class I MHC target cell Antigen presenting cell • The MCH/peptide-TCR interaction is facilitated by the CD4 or CD8 co-receptor. CD8 CD4 CD4+ T cell CD8+ T cell TCR complex TCR complex
Antigen processing: Endogenous pathway All nucleated cells can process endogenous proteins and present fragments on their class I MHC. Display of MHC I + peptide on cell surface degradation Vesicle carrying MHC I-peptide Cytoplasmic proteins Processing in E.R. and complexing with MHC I Endoplasmic reticulum Nucleus
Antigen processing: Exogenous pathway Professional antigen presenting cells ingest microbes and free particles, degrade them in lysozomes, and present fragments to CD4+ T cells on MHC II. Display of MHC II + peptide on cell surface Ingestion of microbe Vesicle fusion, assembly of peptide/MHC II Vesicle carrying MHC II Degradtion in lysozome MHC II is assembled in ER Endoplasmic reticulum Nucleus
CD4+ T cell activation • T cells require co-stimulation for activation -- binding of the TCR to MHC/peptide is not enough to activate a T cell by itself. • B7 on an APC binds to CD28 on the T cell to deliver a co-stimulatory signal. (see figure 13-8). • Activation by peptide/MHC-TCR binding plus a co-stimulatory signal leads to Interleukin-2 (IL-2) release and up-regulation of the IL-2 receptor on the T cell. • IL-2 stimulates growth and proliferation of T cells.
CD8+ T cell activation • A naïve circulating CD8+ T cell also requires co-stimulation to become an “armed” effector cell. • A CD8+ T cell can be activated by an APC displaying MHC I/peptide along with B7 (CD8+ cells also have CD28). • Activation of the CD8+ cell causes upregulation of the IL-2 receptor and production of IL-2, leading to growth and proliferation. • An activated CD8+ T cell can sustain itself on its own IL-2 production, once activated.