Download

1 / 23
240 likes | 477 Views
Actin Filament Assembly, Growth, Branching, Capping and Disassembly are Controlled by Actin Binding Proteins. Actin polymerization is induced by Arp 2/3 protein complex at the surface of Listeria monocytogenes Matthew D. Welch et al.

E N D
Actin Filament Assembly, Growth, Branching, Capping and Disassembly are Controlled by Actin Binding Proteins
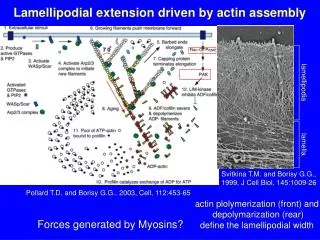
Actin polymerization is induced by Arp 2/3 protein complex at the surface of Listeria monocytogenes Matthew D. Welch et al. The pathogenic bacterium Listeria monocytogenes is capable of directed movement within the cytoplasm of infected host cells. Propulsion is thought to be driven by actin polymerization at the bacterial cell surface and moving bacteria leave in their wake a tail of actin filaments. Actin assembly by L. monocytogenes requires the bacterial surface protein ActA and protein components present in host cell cytoplasm. We have purified an eight-polypeptide complex that possesses the properties of the host-cell actin polymerization factor. The pure complex is sufficient to initiate ActA-dependent actin polymerization at the surface of L. monocytogenes, and is required to mediate actin tail formation and motility. Two subunits of this protein complex are actin-related proteins (ARPs) belonging to the Arp2 and Arp3 subfamilies. The Arp3 subunit localizes to the surface of stationary bacteria and the tails of motile bacteria in tissue culture cells infected with L. monocytogenes; this is consistent with a role for the complex in promoting actin assembly in vivo. The activity and subunit composition of the Arp2/3 complex suggests that it forms a template that nucleates actin polymerization.
Actin Bundling a-actinin and fimbrin cross-link actin filaments
Actin Bundling a-actinin and fimbrin cross-link actin filaments
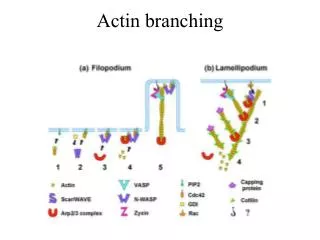
Diversity of Actin Bundling Proteins a-actinin, fimbrin, and villin form parallel cables -stiff connections between actin-binding domains Spectrin and filamin form actin filament webs of gels -flexible connections between actin-binding domains (filamanin) -stiff bent connection (spectrin)
Microvilli (mv) are specialized actin-based structures Increases surface area for Absorption by20-fold Villin and fimbrin cross link 20-30 actin filaments in mv Villin introduced into fibroblasts Induces microvilli formation
Filamin makes 3D Crosslinks
CHI- complex haploinsufficiency and actin haploinsufficiency has been gaining in appreciation as an important influence on human disease human individuals may have as many as 45 CHI gene pairs
Spectrin, glycophorin, and band 3 Correspond to >60% of membrane Proteins in RBCs Spectrin is the most abundant, 25% 250000 copies/cell Principle component of the cytoskeleton If dissociated, the red blood cell fragments into vesicles
Spectrin forms heterodimers And tetramers, and is part of a Junctional complex Ankyrin joins spectrin to the PM by The TM protein band 3 Look how small actin filaments are here! Why? This weblike structure allows cells to deform in capillaries Spectrin mutations cause round and fragile RBCs and anemia
PTEN and polarity: phosphodiesterase and an inhibitor of the phospho-AKT pathway by removing the 3' phosphate group of PI (3,4,5)P3.
Formation of Intercellular Junctions during Wound Healing Mammary epithelial cell monolayer was grown to confluence and mechanically wounded. Cells at the free edge lose their intercellular junctions, form lamellipodia and migrate to fill the empty space. At the last stage of healing, cells from opposite sides of the wound have made contact. Actin (red; rhodamine phalloidin) and occludin (blue, anti-occludin antibody) appear at the newly forming intercellular contacts (arrow). Nuclei were stained green (Syto) to help identify cells.
Reorganizing the cytoskeleton is critical for wound healing Triple label showing microtubules (red), F-actin (green) and myosin-2 (blue) around wound made in a Xenopus oocyte. The microtubules are organized into a radial array that encloses the ring of F-actin and myosin-2B.
Technology: Novel antisense approach provides a rapid spatially and temporally controllable knockdown of specific connexin proteins. A single topical application of Cx43 antisense gel results in a dramatic increase in the rate of wound closure.
Actin depolymerizing factor (ADF)Cofilin binds to Actin It twists the filament, making it easier for subunits at the Minus end of the filament to dissassemble
Importantly, the stimulus-responsive function of ADF/cofilin is regulated by phosphorylation of a single serine residue. In response to stimuli, ADF is dephosphorylated. The stimuli, such as growth factors, chemotactic peptides, or agents increasing the levels of [Ca2+]i and cAMP, promote the reorganization of the actin cytoskeleton. In quiescent cells, ADF/cofilin appears diffusely distributed in the cytoplasm, the activated (dephosphorylated) protein translocates to regions of the cells where actin filaments are highly dynamic like the leading edge of ruffled membranes, the cleavage furrow of dividing cells, or the neuronal growth clone. Dephosphorylation correlates with increased motility and extension of cellular processes (Carlier et al., 1999). ADF/cofilin increases the turnover of actin filaments which powers actin motility.