Download

1 / 28
430 likes | 1.09k Views
RNA Metabolism Transcription and Processing. CH353 April 1, 2008. Types of RNA. Present in all cells, mitochondria and chloroplasts Messenger RNA (mRNA) – encode protein sequences from DNA Transfer RNA (tRNA) – decode mRNA sequences; activate specific amino acids for protein synthesis

E N D
RNA MetabolismTranscription and Processing CH353 April 1, 2008
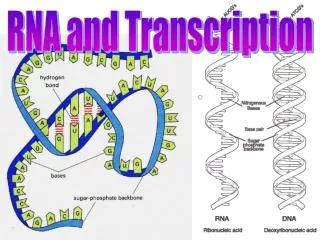
Types of RNA Present in all cells, mitochondria and chloroplasts • Messenger RNA (mRNA) – encode protein sequences from DNA • Transfer RNA (tRNA) – decode mRNA sequences; activate specific amino acids for protein synthesis • Ribosomal RNA (rRNA) – catalytic components of ribosomes; for tRNA binding, codon-anticodon recognition, peptidyl transfer Present predominantly in eukaryotic cells • Catalytic RNAs – components of ribonucleoprotein enzymes • 7S RNA – component of signal recognition particle for secretion • Small nuclear RNAs (snRNAs) – spliceosome subunit components • Small nucleolar RNAs (snoRNAs) – guides for rRNA modification • MicroRNAs (miRNAs) – silencing gene expression
RNA Polymerase Reaction General reaction: (NMP)n + NTP → (NMP)n+1 + PPi Template + n NTP → Template + ppp(NMP)n + n PPi • Requires DNA template, NTP’s and Mg2+ (Initiator NTP is primer) • Reaction driven by hydrolysis of PPi; ∆G’º = -19 kJ/mol Mechanism: nucleophilic attack by 3’-OH on α-phosphate of NTP with PPi as leaving group
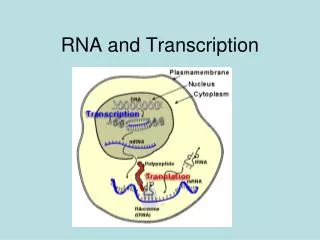
Basic Properties of Transcription • Transcription from DNA without strand separation • Transient transcription bubble of single stranded DNA • Transcription synthesizes single stranded RNA • Transient RNA-DNA hybrid intermediate • Transcription is specific to DNA strand • Template strand DNA transcribed; reverse complement of RNA • Nontemplate or coding strand of DNA; same sense as RNA • Transcription has an initiation site on DNA • RNA synthesis begins at promoters • no primer required: GTP + NTP → (5’)pppGpN-OH(3’) + PPi • Transcription has a termination site • RNA synthesis ends at terminators; defines DNA template
Template and Nontemplate Strands Transcription Units of Adenovirus Genome • Nontemplate sequences same sense as RNA transcripts • Templates for RNA synthesis • do not include entire DNA strand • could be on either or both DNA strands • A sequence and its complement do not both encode proteins
Transcription with E. coli RNA Polymerase Topoisomerase I Topoisomerase II Transcription bubble ~17 bp Rewinding Unwinding Nontemplate strand Template strand RNA-DNA hybrid ~8 bp
Properties of E. coli RNA Polymerase • Large complex (Mr 390,000) of 5 core subunits (a2bb’w) • RNA polymerase holoenzyme has additional s subunit required for specific initiation • Most common is s70 (Mr 70,000) for recognition of most promoters; s32 for heat shock promoters; s54 for regulation by enhancers (NtrC) • RNA polymerases lack 3’ → 5’ exonuclease proofreading activity; can remove incorrect nucleotide by reversal of polymerase reaction • Error rate: 10-4 to 10-5 • RNA polymerases are highly processive; dissociation of polymerase from DNA terminates transcription • RNA polymerization rate: 50 – 90 nucleotides / second (comparable to DNA polymerase II); transcription of most genes in < 1 minute
Promoters for E. coli RNA Polymerase • Recognized by holoenzyme with s70 subunit (most common) • Alignment of RNA start sites reveals upstream consensus sequences • TTGACA at -35 and TATAAT at -10 relative to start (+1) • Strong promoters have consensus sequences and additional A-T rich UP element (between -40 and -60)
Transcription with E. coli RNA Polymerase Initiation and Elongation • E. coli RNA polymerase holoenzyme binds to promoter sequence • Closed complex: RNA polymerase bound but DNA still double stranded • Open complex: 12-15 bp region of DNA from within -10 to +2 or +3 is unwound • Transcription initiation: formation of full transcription bubble; conformational change to elongation form • Elongation form: complex moves away from promoter (promoter clearance); s subunit dissociates after first 8-9 nucleotides are polymerized
Transcription with E. coli RNA Polymerase Termination • Rho (r) independent (shown) • transcription of sequence that can form hairpin loop; • followed by AAA in template for UUU in transcript • pausing of RNA polymerase allows hairpin loop formation and disruption of RNA-DNA • Rho (r) dependent • has CA-rich sequences in template and r binding sites on transcript • the r protein has helicase activity; uses ATP for translocation on RNA
Regulation of E. coli Transcription • Specific s subunits determine promoter selection • for generalized changes, e.g. development stage (sporulation) or stress response (heat shock) • Activation of transcription (Positive Regulation) • cAMP receptor protein (CRP) involved in catabolite activation; increases transcription of gene involved in utilization of carbon sources other than glucose • CRP bound to cAMP binds to DNA upstream of weak promoters • Repression of transcription (Negative Regulation) • lac and trp repressors block transcription by binding to operator sequences within or downstream of promoters • lac repressor bound to inducer doesn’t bind to operator
DNA Footprinting Analysis • Used for qualitative analysis of DNA-protein binding and localizing binding sequences • Uses end-labeled DNA and partial DNase digestion + bound protein • Partially digested DNA is analyzed by denaturing PAGE • Sequences binding to protein are protected from DNase digestion – indicated by missing bands on gel
Electrophoretic Mobility Shift Assay (EMSA) • Used for quantitative analysis of DNA binding proteins • DNA fragment containing a promoter region is labeled • DNA probe incubated with protein fractions suspected of having binding activity • Assayed fractions are analyzed by non-denaturing PAGE • Electrophoretic mobility of DNA is slower if bound to a protein Arrow indicates migration of bound DNA probe EMSA of protein purification fractions Fractions containing TFIIC2 bind labeled VA1 DNA probe, slowing its mobility on non-denaturing PAGE Yoshinaga et al. (1989) J.Biol.Chem 264, 10726
Eukaryotic RNA Polymerases RNA polymerase I II III Subunits 141216 unique abb’w-like 5 5* 5 common 4 4 4 unique 5 3 7 Inhibition [a-amanitin] (resistant)low high Products pre-rRNA mRNAs, tRNAs, (28S, 5.8S, 18S) 5 snRNAs U6 snRNA, 5S rRNA, 7S RNA * large subunit has carboxy terminal domain (CTD)
Properties of RNA Polymerase II • RBP1, largest subunit (Mr 220,000) • is homologous to E. coli RNA polymerase b’ subunit • has unusual carboxyl-terminal domain (CTD) with heptad (-YSPTSPS-) repeats of 27x (yeast) or 52x (human) plus unstructured linker; CTD extends from main polymerase structure ~ 90 to 160 nm • RNA polymerase II promoters • Many have TATA box -35 to -26 bp from start (T-A-T-A-A/T-A-A/T-A/G) • Some have initiator element instead of TATA box (Y-Y-A+1-N-T/A-Y-Y-Y); these also have a 20-50 bp CG-rich region ~100 bp upstream of start • Many have promoter-proximal elements (control regions) within 100-200 upstream of start
Transcription with RNA Polymerase II • Assembly of RNA polymerase and transcription factors at promoter • Formation of closed complex • TFIIH (12 subunits) has helicase, kinase and DNA repair activities • Initiation and promoter clearance • TFIIH phosphorylates CTD of Pol II causing conformation change initiating transcription; TFIIE, TFIIH released after 1st 60-70 nt RNA • Elongation • Elongation factors suppress pausing and coordinate RNA processing; • pTEFb phosphorylates CTD • Termination and Release • Pol II dephosphorylated after release
Structure and Synthesis of mRNA Caps • Most eukaryotic mRNAs have 5’ cap structure • 7-methylguanosine with (+) charge linked to 5’-terminal nucleotide of mRNA by 5’,5’ triphosphate • Unique structure important for translation initiation • Cap formed by transfer of guanylate to 5’ diphosphate and methylation using S-adenosylmethionine
Splicing of mRNA Transcripts Formation of Spliceosome: (5 snRNAs + 50 proteins) • U1 snRNP binds at splice donor site • U2 snRNP binds at branch point site (+ ATP) • U4:U6 snRNP and U5 snRNP bind forming inactive splicesome (+ ATP) • U4 snRNP and U1 snRNP released, U6 snRNP binds splice donor activing spliceosome (+ ATP) • Splicing occurs by 2 step mechanism with lariat intron and spliced exons as products splice donor branch point splice acceptor
Splicing Reaction Mechanism Mechanism for spliceosomal introns and self-splicing group II introns 2 step splicing mechanism: • 2’ OH of adenosine at branch point is nucleophile for attack on splice donor phosphodiester bond; 3’ OH of 5’ exon is leaving group • 3’ OH of 5’ exon (splice donor) is nucleophile for attack on splice acceptor phosphodiester bond; 3’ OH of intron is leaving group Products: spliced RNA + intron RNA with 2’,5’ branch (lariat)
Coupling Transcription and RNA Processing • C-terminal domain (CTD) provides attachment sites for complexes involved with RNA processing • Cap-synthesizing complex binds to CTD and 5’ end of mRNA precursor • 5’ end of RNA is capped • Cap-binding complex binds CTD and 5’ cap of mRNA • Spliceosome components bind CTD, capturing splice donors and branch points of nascent mRNA
Termination and Polyadenylation • Eukaryotic mRNA have a 3’ terminal poly(A) tail (80 – 250 nucleotides) • Polyadenylation linked to transcription termination • Enzyme complex binds CTD cleaving RNA at poly(A) site; between AAUAAA and GU-rich sequences • Polyadenylate polymerase of enzyme complex adds poly(A) to 3’ OH: RNA + nATP → RNA-(pA)n + nPPi
Alternative Polyadenylation Sites Poly(A) site choice forms diversity of 3’ ends for mRNA; C-terminal ends for proteins Alternative Splicing Patterns A splice donor may have multiple splice acceptors, forming diverse mRNAs and encoded proteins Alternative Splicing of mRNA Transcripts
Longer brain transcript is processed at exons 1-2-3-5-6 encoding CGRP (calcitonin gene-related peptide) – a neurotransmitter 1 gene provides 2 different proteins depending on alternate processing of RNA Precursor mRNA has 2 poly(A) sites: one recognized in thyroid and other recognized in brain Shorter thyroid transcript is processed at exons 1-2-3-4 encoding calcitonin – a calcium regulating hormone Splicing of Calcitonin/CGRP mRNA
Processing of Human rRNA Precursors In the nucleolus: • RNA pol I transcribes 45S pre-rRNA from multiple rRNA genes • Small nucleolar RNAs (snoRNA) guide methylation and cleavage of the precursor into mature 18S, 5.8S and 28S rRNAs • >100 of 14000 nucleotides are methylated
E. coli RNA polymerase transcribes RNA from one of 7 rRNA genes Each precursor contains 16S, 23S and 5S rRNAs plus 1 – 2 tRNAs Site-specific methylation of pre-rRNA using guide RNAs Nuclease cleavage with: RNase III RNase P RNase E Processing with various specific nucleases Processing of Bacterial rRNA Precursors
Processing tRNA Precursors • Yeast tRNA is processed from precursor by removing the 5’ end with RNase P (a ribozyme), then the 3’ end with RNase D • The terminal CCA(3’) is added 1 nucleotide at a time by the enzyme tRNA nucleotidyl transferase – template independent RNA synthesis • Bases are modified during the processing of tRNA • Some tRNAs are spliced, removing the intron in the last step