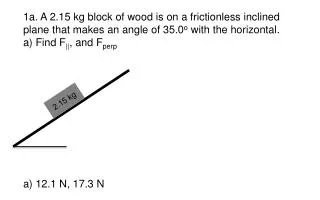
Download

1 / 37
380 likes | 625 Views
Buněčná membrána a její modelování. Josef Rosenberg 28.2.2012 Přednáška v rámci projektu Rozvoj výzkumných kapacit v oblasti polymerních membrán (projekt OP VK - CZ.1.07/2.3.00/20.0107). BUNĚČNÁ MEMBRÁNA A JEJÍ MODELOVÁNÍ. Úvod, motivace Buněčná membrána – struktura Integrální proteiny

E N D
Buněčná membrána a její modelování Josef Rosenberg 28.2.2012 Přednáška v rámci projektu Rozvoj výzkumných kapacit v oblasti polymerních membrán (projekt OP VK - CZ.1.07/2.3.00/20.0107)
BUNĚČNÁ MEMBRÁNA A JEJÍ MODELOVÁNÍ • Úvod, motivace • Buněčná membrána – struktura • Integrální proteiny • Membrána buňky hladkého svalu • Matematické modely přestupů • Intersticiální buňka Cajalova • Shrnutí, závěr
BUNĚČNÁ MEBRÁNA A JEJÍ MODELOVÁNÍ • Proč se (bio)mechanik zabývá buněčnou membránou? • Všechny makromechanické procesy jsou řízeny na úrovni buněk • Buňka (i některé organely) je vždy chráněna membránou, která ale sehrává velmi důležitou roly v klíčových procesech!
ÚVOD, MOTIVACE • V řízení procesů, jimiž se biomechanika zabývá, buněčná membránahrajedůležitou roli . • Je tomu tak • při řízení svalové kontrakce cestou změny koncentrace vápníkových iontů ve svalové buňce, • při vzniku akčního potenciálu – pacemakerové buňky, • při přenosu signalu mezi axonem a svalovou buňkou, • při vlastním přenosu akčního potenciálu nervovým vláknem. • Membrána komunikuje s vnitrobuněčnými organelami (ER, mitochondrie atp.)
BUNĚČNÁ MEMBRÁNA - STRUKTURA • Základem membrány je fosfolipidová dvojvrstva. • Fosfolipidy jsou tvořeny hydrofilními (vodu přitahujícími, polárními) hlavičkami a hydrofobními (vodu odpuzujícími, nepolárními) ocasy. Fosfátové hlavičky tvoří vnější a vnitřní povrch. Ocasy jsou tvořeny mastnými kyselinami a tvoří vnitřní isolační vrstvu.
BUNĚČNÁ MEMBRÁNA - STRUKTURA • Další důležitou komponentou membrán jsou proteiny. Tvoří zhruba 50% hmoty membrány. Dělí se na tzv. integrální, jež prostupují celou fosfolipidovou vrstvu, a periferní, jež jsou uchyceny pouze na vnitřní nebo vnější straně membrány. Integrální protein se podílí na transportu látek membranous, vnější periferní protein často působí jako receptory, jež vážou hormony či jiné látky. Tato vazba pak ovlivňuje procesy v buňce. Rovněž se podílí na rozpoznávání buněk a na dalších jevech. Vnitřní periferní proteiny jsou často připojeny na cytoskeleton a podílí se na zpevnění buňky. Mohou to být rovněž enzymy katalyzující reakce v cytoplazmě . • Třetí významnou složkou buněčné membrány jsou uhlovodany (carbohydrates). Nalézají se na vnějším povrchu vázané buď na protein (pak se nazývají glykoproteiny) či přímo na fosfolipid a pak hovoříme o glykolipidech. Jsou to krátké polysacharidy a tvoří buněčný obal – gykokalyx – sloužící k ochraně a rozpoznávání buněk.
BUNĚČNÁ MEMBRÁNA - STRUKTURA • Prostup látek membránou lze rozdělit do několika forem: • Pasivní transport, který zahrnuje • Difúze lipidovou dvojvrstvou • Difúze iontovými kanály • Facilitovaná difúze • Osmóza • Aktivní transport • Exocytóza a endocytóza
INTEGRÁLNÍ PROTEINY • Pasivní transport Pasivní transport je realizován integrálními proteiny tvořícími buď kanál nebo nosič ,jenž periodicky otevírá a zavírá vrátka na vnější a vnitřní straně membrány. V obou případech jsou tyto proteiny vždy specifické pro daný typ molekuly. Jejich otevření či zavření může být buňkou řízeno. Směr je dán pouze koncentračním gradientem.
INTEGRÁLNÍ PROTEINY DRASLÍKOVÝ KANÁL (OTEVŘENÝ)
INTEGRÁLNÍ PROTEINY • Difúze iontovými kanály Tento typ pasivního transportu je realizován integrálními proteiny tvořícími transmembránový kanál s centrálním pórem. Kanál může být stále otevřený nebo se může zavírat či otevírat vrátky v závislosti na aktuální situaci (elektrické napětí, mechanická tenze, přítomnost ligandu). Kanály mohou být více či méně specifické pro určitý typ iontu (např. sodíkové kanály, draslíkové kanály, nespecifické kationtové kanály). Směr pohybu iontu je určován elektrochemickým gradientem (koncentrační i elektrický). • Facilitovaná difúze Transport probíhá ve směru elektrochemického gradientu za pomoci integrálních proteinů – nosičů, specifických pro určitý typ molekuly. Transport je spojený se změnou konformace proteinu a vykazuje jistou limitní maximální rychlost transportu (všechny nosiče jsou při daném gradientu plně vytíženy a další zvyšování gradientu nemůže transportní rychlost zvýšit). Příkladem může být transport glukózy
INTEGRÁLNÍ PROTEINY • Osmoza Osmoza je difuze vodních molekul lipidovou dvojvrstvou nebo prostřednictvím vodních kanálů, akvaporinů. Směr a intenzita osmozy je dána rozdílem koncentrace molekul vody na obou stranách membrány. Čím je vodní roztok nějaké látky koncentrovanější, tím je menší podíl molekul vody a i nižší tzv.vodní potenciál . Pro vodu bez příměsi je , pro roztoky nabývá záporných hodnot (nikdy není kladný). Dříve používaný termín osmotický tlak se liší od vodního potenciálu znaménkem.
INTEGRÁLNÍ PROTEINY • Difúze lipidovou dvojvrstvou Jen málo látek může prostupovat buněčnou membránou pouze vlivem jejich koncentračního gradientu. Mohou to být látky rozpustné v lipidech jako jsou steroidy nebo velmi malé molekuly jako jsou molekuly vody, kyslíku či oxydu uhličitého. Tento prostup není nijak kontrolován.
INTEGRÁLNÍ PROTEINY • Aktivní transport Aktivní transport se děje integrálními proteiny – proteinovými pumpami. Tyto proteiny jsou zároveň enzymy ATP. Katalyzují hydrolyzu ATP a uvolněnou energii využívají ke změně konformace vedoucí k přenosu příslušné molekuly proti elektrochemickému gradientu
INTEGRÁLNÍ PROTEINY • Jako příklad uveďme Na+/K+ pumpu,která patří mezi nejčastější a nejdůležitější. Tato pumpa transportuje během jednoho cyklu tři ionty sodíku ven z buňky a dva ionty draslíku do buňky. Na tento transport je spotřebována energie hydrolýzy jedné molekuly ATP. Tím vytváří koncentrační gradient pro ionty sodíku (vysoká koncentrace vně buňky, nízká uvnitř) a draslíku (vysoká koncentrace uvnitř buňky, nízká vně).
INTEGRÁLNÍ PROTEINY • Exocytóza a endocytóza • Předchozí procesy dovolují prostup pouze malých molekul. Velké molekuly mohou procházet membránou v tzv. vesiklech – viz obr. Hovoříme o endocytose, kdy se jedná o transport do buňky a o exocytose, jedná-li se o transport z buňky ven. • U endocytosy rozlišujeme dle velikosti transportovaných molekul či dokonce buněk tzv. pinocytosu (malé molekuly,“buněčné pití“) či fagocytosu (velké molekuly či buňky).
MATEMATICKÉ MODELY PŘESTUPŮ • Rychlost změny koncentrace kalcia v cytoplazmě je dána • Uvolňováním Ca2+ z IP3 – senzitivních zásobáren Poznamenejme zde, že Parathimos (2007) uvádí pro tento jev vztah • Na vápníku závislým uvolněním Ca2+ z ER/SR (CICR) prostřednictvím ryonidinových receptorů/kanálů Fakulta aplikovaných věd Katedra mechaniky
MATEMATICKÉ MODELY PŘESTUPŮ • tokem vápníku povrchovou membránou (VOC – voltage operated chanells, ICaL) • nespecifickým tokem/únikem kalcia z ER/SR • tokem vápníku povrchovou membránou mechanosenzitvními kanály (kanály řízenými mechanickou deformací buňky – SAC) Fakulta aplikovaných věd Katedra mechaniky
MATEMATICKÉ MODELY PŘESTUPŮ • Tokem vápníku z buňky povrchovou membránou Na+-Ca2+ výměníkem • odčerpáváním vápníku do ER/SR prostřednictvím retikulární vápníkové pumpy (SERCA) • odčerpáváním vápníku sarkolemální vápníkovou pumpou ( energeticky zásobované Ca2+ ATPasou ) Fakulta aplikovaných věd Katedra mechaniky
MATEMATICKÉ MODELY PŘESTUPŮ • SERCA pumpa musí dopravit ionty vápníku oproti značnému koncentračnímu gradientu – uvnitř buňky je typická koncentrace Ca2+ řádově 0.1 , zatím co uvnitř sarkoplazmatického reticula je to 0.1 až 1 mM (M je mol/l) . Energie pro toto čerpání je získávána ze štěpení ATP (odtud název SERCA: „Sarco-Endoplasmic Reticulum Ca2+ ATPasa“). Detailními měřeními bylo zjištěno, že tok této pumpy závisí na koncentraci vápníku uvnitř buňky . • Nejjednoduší model této závislosti vychází z představy, že existují dva stavy pumpy – aktivní A a neaktivní I, přičemž k transportu vápníku dochází pouze v aktivním stavu a rovnováha obou stavu nastává velmi rychle. Jedná se o typický molekulární proces. Označíme-li koncentraci aktivovovaných stavů a neaktivovaných stavů [I], [A] , pak pro rychlosti přechodu ze stavu A do stavu I a naopak platí
MATEMATICKÉ MODELY PŘESTUPŮ • Podmínka rovnováhy pak má tvar • Z experimentů bylo zjištěno, že tato konstanta je nepřímo úměrná kvadrátu koncentrace vápníku v buňce a tedy • Uvážíme-li, že platí ,pak lze odvodit vztah pro sigmoidální závislost koncentrace aktivovaného stavu SERCA pump na koncentraci intracelulárního vápníku • jenž odpovídá vztahu pro v Koenigsbergově modelu.
MATEMATICKÉ MODELY PŘESTUPŮ • VOCC • Jako další příklad uveďme odvození vztahu pro tok vápníku napěťově řízenými kanály typu L. Vyjdeme z práce (Burzstyn, 2007). Tento tok je dán vztahem • kde je vodivost kanálů L-typu na buněčné membráně, je Nernstův potenciál daný koncentracemi iontů vně a uvnitř membrány, je valence vápníku, objem buňky a F Faradayova konstanta.
MATEMATICKÉ MODELY PŘESTUPŮ • Pro každý typ iontu existuje tzv. reverzní potenciál (rovnovážný potenciál)E, kdy celkový tok tohoto iontu membránou je nulový. Tok vyvolaný elektrickým polem se ruší s tokem vyvolaným rozdílem koncentrací. Platí pro něj Nernstova rovnice
MATEMATICKÉ MODELY PŘESTUPŮ • Lze však určit membránový potenciál (klidový potenciál), pro který součet všech toků je nulový. Je dán tzv. Goldmanovou rovnicí . Pro tři monovalentní ionty draslíku, sodíku a chloru např. má tvar
MATEMATICKÉ MODELY PŘESTUPŮ • Závislost vodivosti na potenciálu lze popsat aktivační křivkou Boltzmanova typu je funkce potenciálu • Po postupném dosazení obdržíme konečný vztah • Porovnáme-li tento vztah se vztahem uvedeným výše (viz (Koenigsberger,2004)), vidíme, že zde je Nernstův potenciál uvažován jako konstantní
CAJALOVA BUŇKA • Beridge:
CAJALOVA BUŇKA • Cajalova intersticiální buňka • Různé modely: Sauders – pacemaker unit • Beridge – oslabená role mitochondrií
CAJALOVA BUŇKA R. A. Faville,* A. J. Pullan, K. M. Sanders, S. D. Koh, C. M. Lloyd, and N. P. Smith • Biophysically Based Mathematical Modeling of Interstitial Cells of Cajal Slow Wave Activity Generated from a Discrete Unitary Potential Basis Biophysical Journal Volume 96 June 2009 4834–4852 Pacemakerová jednotkat:
CAJALOVA BUŇKA • Pacemakerová jednotka:
CAJALOVA BUŇKA Alberto Corrias, M.L.Buist: • Ca-cyto • Vm • Ca-m • Ca-ER
SHRNUTÍ, ZÁVĚR • Buněčná membrána je nesmírně složitá „smart“ struktura • Procesy, jichž se aktivně zúčastňuje, lze popsat nelineárními dynamickými systémy a pro jejich analýzu využít příslušné metody • Ač je buněčná membrána značně probádaná, řada procesů je ještě neznámých • I u procesů známých často chybí matematický model Vděčný objekt pro biomechaniky
NERVOVÁ BUŇKA • Šíření akčního potenciálu Speciálním typem buňky jsou neurony , jež hrají klíčovou roli při ecxitaci svalových buněk. Neuron má obvykle několik výběžků nazývaných dendrity ,které vystupují z těla buňky a větví se, a jeden dlouhý vláknitý axon (neurit) vystupující z axonového hrbolku ( axon hillock) . Zatím co dendritů bývá větší počet , neurony zpravidla obsahují jeden axon, kterým se šíří akční potenciál. Axon se povětšině na konci dělí do velkého počtu větví zakončených nervovými (synaptickými) zakončeními (knoflíky synaptic knobs -), která jsou napojena buď na dendrity či těla dalších neuronů nebo efektorové buňky. Tato spojení se nazývají synapse.