Download

1 / 54
540 likes | 554 Views
Learn about sequence-specific activators, transcription factors, and their regulatory roles in gene expression. Explore the principles of DNA-binding, classification of transcription factors, and mechanisms of sequence recognition.

E N D
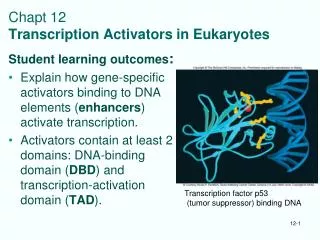
The sequence specific activators: transcription factors • Modular design with a minimum of two functional domains • 1. DBD - DNA-binding domain • 2. TAD - transactivation domain • DBD: several structural motifs classification into TF-families • TAD - a few different types • Three classical categories • Acidic domains (Gal4p, steroid receptor) • Glutamine-rich domains (Sp1) • Proline- rich domains (CTF/NF1) • Mutational analyses - bulky hydrophobic more important than acidic • Unstructured in free state - 3D in contact with target? • Most TFs more complex • Regulatory domains, ligand binding domains etc DBD N TAD C
TF classification based on structure of DBD • Two levels of recognition • 1. Shape recognition • Anhelix fits into the major groove in B-DNA. This is used in most interactions • 2. Chemical recognition • Negatively charged sugar-phosphate chain involved in electrostatic interactions • Hydrogen-bonding is crucial for sequence recognition bHelix-Loop-Helix (Max) Zinc finger Leucine zipper (Gcn4p) p53 DBD NFkB STAT dimer
Alternative classification of TFs on the basis of their regulatory role • Classification questions • Is the factor constitutive active or requires a signal for activation? • Does the factor, once synthesized, automatically enter the nucleus to act in transcription? • If the factor requires a signal to become active in transcriptional regulation, what is the nature of that signal? • Classification system • I. Constitutive active nuclear factors • II. Regulatory transcription factors • Developmental TFs • Signal dependent • Steroid receptors • Internal signals • Cell surface receptor controlled • Nuclear • Cytoplasmic
Classification - regulatory function Brivanlou and Darnell (2002) Science295, 813 -
Sequence specific DNA-binding- essential for activators • TFs create nucleation sites in promoters for activation complexes • Sequence specific DNA-binding crucial role
How is a sequence (cis-element) recognized from the outside? Shape recognition Chemical recognition Electrostatic interaction Form/ geometry Hydrogen- bonds Hydrophobic interaction
Complementary forms The dimension of anhelix fits the dimensions of the major groove in B-DNA Side chains point outwards and are ideally positioned to engage in hydrogen bonds
Direct reading of DNA-sequenceRecognition of form • The dimension of an a-helix fits the dimensions of the major groove in B-DNA • Most common type of interaction • Usually multiple domains participate in recognition • dimers of same motif • tandem repeated motif • Interaction of two different motifs • recognition: detailed fit of complementary surfaces • Hydration /water participates • seq specific variation of DNA-structure
Example • Steroid receptor
How is a sequence (cis-element) recognized from the outside? Shape recognition Chemical recognition Electrostatic interaction Form/ geometry Hydrogen- bonds Hydrophobic interaction
Next level: chemical recognition - reading of sequence information • Negatively charged sugar-phosphate chain = basis for electrostatic interaction • Equal everywhere - no sequence-recognition • Still a main contributor to the strength of binding
How is a sequence (cis-element) recognized from the outside? Shape recognition Chemical recognition Electrostatic interaction Form/ geometry Hydrogen- bonds Hydrophobic interaction
D A A Recognition by Hydrogen bonding • Hydrogen-bonding is a key element in sequence specific recognition • 10-20 x in contact surface • Base pairing not exhausted in duplex DNA, free positions point outwards in the major groove
Docked prot side chains exploit the H-bonding possibilities for interaction • Hydrogen-bonding is essential for sequence specific recognition • 10-20 x in contact interphase • Most contacts in major groove • Purines most important • A Zif example
Interaction: Protein side chain - DNA bp • Close up • Amino acid side chains points outwards from the a-helix and are optimally positioned for base-interaction
How is a sequence (cis-element) recognized from the outside? Shape recognition Chemical recognition Electrostatic interaction Form/ geometry Hydrogen- bonds Hydrophobic interaction
The Homeodomain-family: common DBD-structure • Homeotic genes - biology • Regulation of Drosophila development • Striking phenotypes of mutants – body-parts move • Control genetic developmental program • Homeobox / homeodomain • Conserved DNA-sequence “homeobox” in a large number of genes • Encode a 60 aa “homeodomain” • A stably folded structure that binds DNA • Similarity with prokaryotic helix-turn-helix • 3D-structure determined for several HDs • Drosophila Antennapedia HD (NMR) • Drosophila Engrailed HD-DNA complex (crystal) • Yeast MAT2
Homeodomain-family: common DBD-structure • Major groove contact via a 3 -helix structure • helix 3 enters major groove (“recognition helix”) • helix 1+2 antiparallel across helix 3 • 16 -helical aa conserved • 9 in hydrophobic core • some in DNA-contact interphase (common docking mechanism?)
Homeodomain-family: common DBD-structure • Minor groove contacted via N-terminal flexible arm • R3 and R5 in engrailed and R7 in MAT2 contact AT in minor groove • R5 conserved in 97% of HDs • Deletions and mutants impair DNA-binding • Loop between helix 1 and 2 determines Ubxversus Antp function • Close to DNA • exposed for protein-protein interaction
HD-paradox: what determines sequence specificity? • Drosophila Ultrabithorax (Ubx), Antennapedia (Antp), Deformed (Dfd) and Sex combs reduced (Scr): closely similar HD, biological role very different • Minor differences in DNA-binding in vitro • TAAT-motif bound by most HD-factors • contrast between promiscuity in vitro and specific effects in vivo • Swaps reveal that surprisingly much of the specificity is determined by the N-terminal arm which contacts the minor groove • Swaps: Antp with Scr-type N-term arm shows Scr-type specificity in vivo • Swaps: Dfd with Ubx-type N-term arm shows Ubx-type specificity in vivo • N-terminal arm more divergent than the rest of HD • R5 and R7 (contacting DNA) are present in both Ubx, Antp, Dfd, and Scr • Other tail aa diverge much more
Solutions of the paradox • Conformational effects mediated by N-term arm • Even if the -helical HDs are very similar, a much larger diversity is found in the N-terminal arms that contact the minor groove • Protein-protein interaction with other TFs through the N-terminal arm - enhanced affinity/specificity - the basis of combinatorial control • MAT2 interaction with MCM1 - cooperative interactions • Ultrabithorax- Extradenticle in Drosophila • Hox-Pbx1 in mammals
Combinatorial TFs give enhanced specificity • TFs encoded by the the homeotic (Hox) genes govern the choice between alternative developmental pathways along the anterior–posterior axis. • Hox proteins, such as Drosophila Ultrabithorax, have low DNA-binding specificity by themselves but gain affinity and specificity when they bind together with the homeoprotein Extradenticle (or Pbx1 in mammals).
a b N-tail in protein-protein interaction- adopt different conformations HD HD Conformation determined by prot-prot interaction Mat-a2/Mcm-1
It works impressively well • Hox genes
POU-family: common DBD-structure • The POU-name : • Pit-1 pituitary specific TF • Oct-1 and Oct-2 lymphoid TFs • Unc86 TF that regulates neuronal development in C.elegans • A bipartite160 aa homeodomain-related DBD • a POU-type HD subdomain (C-terminally located) • et POU-specific subdomain (N-terminally located) • Coupled by a variable linker (15-30 aa) • POU is a structurally bipartite motif that arose by the fusion of genes encoding two different types of DNA-binding domain.
POU: Two independent subdomains • POUHD subdomain • 60 aa closely similar to the classical HD • Only weakly DNA-binding by itself (<HD) • contacts 3´-half site (Oct-1: ATGCAAAT) • docking similar to engrailed. Antp etc • Main contribution to non-specific backbone contacts • POUspec subdomain • 75 aa POU-specific domain • enhances DNA-affinity 1000x • contacts 5´-half site (Oct-1: ATGCAAAT) • contacts opposite side of DNA relative to HD • structure similar to prokaryotic - and 434-repressors • The two-part DNA-binding domain partially encircles the DNA.
Flexible DNA-recognition • POU-domains have intrinsic conformational flexibility • and this feature appears to confer functional diversity in DNA-recognition • The subdomains are able to assume a variety of conformations, dependent on the DNA element.
Zinc finger proteins • Zinc finger proteins were first discovered as transcription factors. • Zinc finger proteins are among the most abundant proteins in eukaryotic genomes. • Their functions are extraordinarily diverse • include DNA recognition, • RNA packaging, • transcriptional activation, • regulation of apoptosis, • protein folding and assembly, and • lipid binding. • Zinc finger structures are as diverse as their functions.
Examples C4-type Zif fra GATA-1 C2H2-type Zif fra Sp1 (3.fngr) Zn++ LIM-domain type Zif fra ACRP PKC-type Zif
++ ++ Zn Zn ++ ++ Zn Zn ++ ++ ++ ++ Zn Zn Zn Zn Classical TFIIIA-related zinc fingers: n x [Zn-C2H2] • History: Xenopus TFIIIA the first isolated and cloned eukaryotic TF • Function: activation of 5S RNA transcription (RNAPIII) • Rich source : accumulated in immature Xenopus oocyttes as “storage particles” = TFIIIA+5S RNA (≈ 15% of total soluble protein) • Purified 1980, cloned in 1984 • Mr= 38 600, 344 aa • Primary structure TFIIIA • Composed of repeats: 9x 30aa minidomains + 70aa unique region C-trm • Each minidomain conserved pattern of 2Cys+2His • Hypothesis: each minidomain structured around a coordinated zinc ion (confirmed later)
Zinc finger proteins • Finger-like in 2D • Not in 3D
Common features of TFIIIA-related zinc fingers • Consensus for each finger: FXCX2-5CX3FX5FX2HX2-5H • Number of fingers in related factors varies: 2-37 • Number of members exceptionally high • S.cerevisiae genome: 34 C2H2 zinc fingers • C.elegans genome 68 C2H2 zinc fingers • Drosophila genome 234 C2H2 zinc fingers • Humane genome 564C2H2 zinc fingers, (135C3HC4 zinc finger) • We now recognize the classical C2H2 zinc finger as the first member of a rapidly expanding family of zinc-binding modules.
3D structure of the classical C2H2-type of zinc fingers • Each finger = a minidomain with -structure • each finger an independent module • Several fingers linked together by flexible linkers • First 3D structure: the 3-finger Zif268 (mouse) • DNA interaction in Zif268 • major groove contact through -helix in • recognition of base triplets • aa in three positions responsible for sequence recognition: -1, 3 and 6 (rel. til -helix) • Simple one-to-one pattern (contact aa - baser) can a recognition code be defined ?? • DNA interaction in GLI and TTK differs • different phosphate contact • distortion of DNA • finger 1 without DNA contact D N A
The Zif268 prototype • Finger 2 from Zif268 • including the two cysteine side chains and two histidine side chains that coordinate the zinc ion • DNA-recognition residues • indicated by the numbers identifying their position relative to the start of the recognition helix
Finger 3 DNA Finger 2 Finger 1 LINKER Three fingers in Zif268 • Zif268 - first multi-finger structure • recognition of base triplets
Recognition code? • The DNA sequence of the Zif268 site is color coded to indicate base contacts made by each finger.
Structure of the six-finger TFIIIA–DNA complex • In a multi-finger protein some fingers contact base pairs and some will not, but rather function as bridges • Fingers 1–2–3, separated by typical linkers, wrap smoothly around the major groove like those of Zif268 • In contrast, fingers 4–5–6 form an open, extended structure running along one side of the DNA. Of these, only finger 5 makes contacts with bases in the major groove. The flanking fingers, 4 and 6, appear to serve primarily as spacer elements.
Nuclear receptors: 2x[Zn-C4]: • Large family where DBD binds two Zn++ through a tetraedrical pattern of Cys • conserved DBD 70-80 aa • Protein structure • Two “zinc fingers” constitute one separate domain • Two -helices with C3-Zn-C4 N-terminally • These perpendicular on the top of each other with hydrophobic interactions • Mediates trx response to complex extra cellular signals • Evolutionary coupled to multi cellular organisms • Yeast = 0 but C.elegans 233 or 1.5% of genes !! • Sequence prediction: 90% with nuclear receptor DBD has potential ligand-BD • Implies that lipophilic signal molecules have been important to establish communication between cells
Nuclear receptors - DNA interaction • 3D Prot-DNA structure • glucocorticoid receptor + estrogen receptor • Dimer in complex (monomer in solution) • DNA interaction • First “finger” binds DNA • Second “finger” involved in dimerization • Binds to neighboring “major grooves” on same side of DNA • Extensive phosphate contact and recognition helix docked into the groove • specificity determined by 3 aa (E2, G3, A6) in recognition helix • Structured dimer interphase formed upon DNA-binding