Download

1 / 19
190 likes | 491 Views
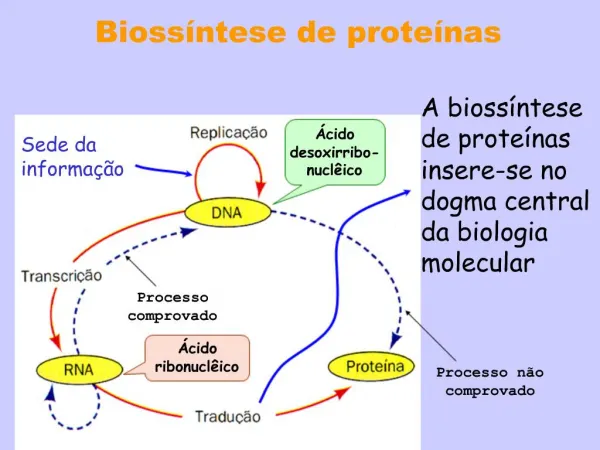
13 SÍNTESIS DE PROTEÍNAS : TRADUCCIÓN. 1. TRADUCCIÓN: DESCODIFICACIÓN DEL ARNm. Etapa en la cual se descodifica el mensaje contenido en el ARNm formado durante la transcripción cuyo mensaje dará lugar a una proteína.

E N D
13 SÍNTESIS DE PROTEÍNAS : TRADUCCIÓN.
1. TRADUCCIÓN: DESCODIFICACIÓN DEL ARNm Etapa en la cual se descodifica el mensaje contenido en el ARNm formado durante la transcripción cuyo mensaje dará lugar a una proteína. Debe pasar del idioma del ácido nucleico (mensaje contenido en cuatro letras) al idioma de las proteínas (con 20 letras) EL CÓDIGO GENÉTICO: establece la correspondencia entre nucleótidos y aminoácidos: los aminoácidos están codificados por tres letras, que son tripletes de bases del ARNm o también denominados codones.
CRACTERÍSTICAS DEL CÓDIGO GENÉTICO. • El código es degenerado: es decir, que es redundante: ya que existen 61 codones que codifican 20 aminoácidos, algún aminoácido debe estar codificado por dos o más tripletes distintos, es decir, hay codones que significan lo mismo es decir, existen codones sinónimos. • El código es universal: vale para todos los organismos conocidos y no es ambiguo, cada codón codifica siempre el mismo aminoácido. • Todos los tripletes de bases tienen sentido y se leen en el ARNm de izquierda a derecha, en sentido a 3’. • El código carece de solapamiento, ningún triplete comparte sus bases con otro triplete; es unidireccional, se lee sólo en sentido a 3’.
Para que los aminoácidos reconozcan a sus tripletes necesitan de un intérprete que hacen de intermediario entre aminoácido y proteína: son los ARNt. CADA MOLÉCULA DE ARNt ESTÁ COMPUESTO DE DOS REGIONES : LA SECUENCIA CCA DEL EXTREMO 3’ Y EL BUCLE DEL ANTICODÓN.
Cada ARNt se une con un único aminoácido: la enzima aminoacil ARNt sintetasa actúa de adaptador con dos sitios específicos: uno reconoce al aminoácido y otro que identifica secuencias de bases características de cada ARNt. Esta enzima cataliza el enlace entre el carboxilo del aminoácido con el hidroxilo del ribonucleótido de adenina del extremo 3’ del ARNt (CCA). • Los ARNt con sus aminoácidos reconocen y se unen mediante su anticodón con los correspondientes tripletes o codones del ARNm. • El ribosoma es capaz de mantener en su interior el ARNm, a los ARNt con sus aminoácidos y forma el enlace peptídicos entre los aminoácidos.
BALANCEO: apareamiento un tanto defectuoso de la tercera base del anticodón y es la causa de la degeneración del códigogenético. La complementariedad entre el codón y el anticodón sólo es máxima en lo que se refiere a las dos primeras bases del anticodón, mientras que la tercera base puede aparearse no sólo con su base complementaria normal, sino también con otras distintas. El balanceo posibilita que un ARNt pueda unirse con su anticodón a codones distintos del mismo aminoácido y también que puede existir más de un ARNt distinto para ciertos aminoácidos. Denominamos ISOACEPTORES a los diferentes ARNt con anticodones distintos que aceptan al mismo aminoácido.
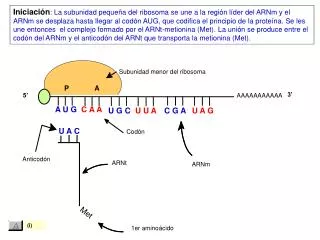
TRADUCCIÓN EN EUCARIOTAS 1. INICIACIÓN DE LA SÍNTESIS DE LA CADENA PEPTÍDICA - Construcción del complejo de iniciación 80 S : formado por un ribosoma unido al ARNm y ARNt iniciador cargado con el aminoácido metionina, ARNt iMet . FI (factores de iniciación) + E de GTP provocan unión de subunidad 40S con el ARNt iniciador, cargado con el aminoácido metionina. Otros FI facilitan que reconozca la “caperuza” de metilguanosina trifosfato y se una con el ARNm en el extremo 5’.
La subunidad pequeña se desplaza 5’ a 3’ con E de ATP y rastrea secuencia hasta el primer codón de iniciación AUG. Se produce la unión del ARNt iniciador con la Met el cual posee el anticodón UAC. 4. Al encajar el ARNtiMet en el codón AUG, se liberan los FI y dejan paso a la subunidad mayor del ribosoma que se acopla con la pequeña, el ARNm y el ARNtiMet formando el complejo de iniciación 80S . En la subunidad mayor distinguiremos tres hendiduras: A,P y E.
2. ELONGACIÓN DE LA CADENA PEPTÍDICA PRIMERA FASE: el sitio P está ocupado por el ARNtiMet y los sitios A y E están vacios. En el sitio A entra el siguiendo ARNt con su aminoácido. Interviene el FE-1 y E del GTP.
SEGUNDA FASE: formación del enlace peptídico. Entre la metionina y siguiente aminoácido. El complejo enzimático peptidiltransferasa debe de estar ayudado por el ARNr 28S asentado en la subunidad grande. Metionina se separa del ARNt y se asocia por enlace peptíco al grupo amino del aminoácido en el sitio A. Obtenemos un dipéptido.
TERCERA FASE: O TRASLOCACIÓN. Entra en juego el FE-2 que con E del GTP obliga al ribo a desplazarse tres nucleótidos sentido 5’ a 3’: primero se desplaza la subunidad grande del cromosoma y al moverse el nuevo codón queda en el sitio A , luego se mueve la subunidad pequeña. El complejo peptidil-ARNt-ARNm queda ahora localizado en el sitio P. El ARNti que era de la metionina pasa al sitio E. FASES SUCESIVAS: al entrar el tercer ARNt con su aminoácido provoca la salida del ARNt del sitio E y se repiten las tres fases anteriores. Así se va produciendo la elongación a un ritmo de 3 a 5 aminoácidos por segundo. Hasta que se incorpore el último aminoácido a la cadena polipeptídica.
3. TERMINACIÓN DE LA SÍNTESIS. La síntesis termina cuando tras una traslocación aparece en el sitio A uno de los tres codones de terminación: UAA, UAG O UGA. Cuando llega este momento un factor proteico de terminación (RF) se une al codón de terminación e impide que algún aminoacil-ARNt se aloje en el sitio A.
4. MADURACIÓN POSTRADUCCIONAL DE LAS PROTEINAS • Al finalizar la traducción los polipéptidos no son funcionales y deben sufrir determinadas modificaciones: • Formación de puentes disulfuropara el plegamiento de las cadenas peptídicas y asociación con otras cadenas para formar estructuras cuaternarias • Adición de grupos prostéticos • Modificaciones covalentes de ciertos aminoácidos ( fosforilaciones, metilaciones,…) • Cortes proteolíticos acortando el péptido. Pérdida de la metionina inicial o eliminación de un péptido intermedio, …
PLEGAMIENTO POSTRADUCCIONAL: CHAPERONAS El plegamiento es espontáneo, a la vez que se van sintetizando, la propia cadena es la que indica como se debe plegar adoptando la conformación espacial adecuada para su funcionamiento biológico adecuado. Hay veces que no es espontáneo y requieren la actuación delas chaperonas moleculares: que son un grupo de proteínas que dan un entorno seguro para que puedan plegarse las cadenas (las chaperonas serán de unos 60.000 Dalton) Toda célula posee una cierta cantidad de chaperonas para que el plegamiento y auto- ensamblaje sea correcto..
También intervienen en el tráfico de proteínas entre citosol y orgánulos celulares (desenrollan para pasar por poros y después vuelven a enrollar) A veces pueden funcionar como proteínas de choque térmico o anti-estrés: actúan como defensa y protección: atenúan condiciones adversas como cambio de temperatura, ausencia de oxígeno,…, evitando así la desnaturalización de las proteínas formadas.
EXPORTACIÓN Y DESTINO Los ribosomas dependiendo del destino que tendrá la proteína estarán localizados en un sitio ó en otro. RIBOSOMAS LIBRES: sintetizan proteínas para procesos metabólicos y aquellas que van al núcleo, mitocondrias, cloroplastos o peroxisomas. RIBOSOMAS DE RER: proteínas para estructuras celulares, hormonas para enviar al exterior de célula o interior de lisosomas.
IMP: en el extremo N—terminal de la proteína se forma un péptido señal de unos 20 ó 30 aminoácidos que será la señal que indicará donde debe ir esa proteína.