Download
1 / 73
730 likes | 860 Views
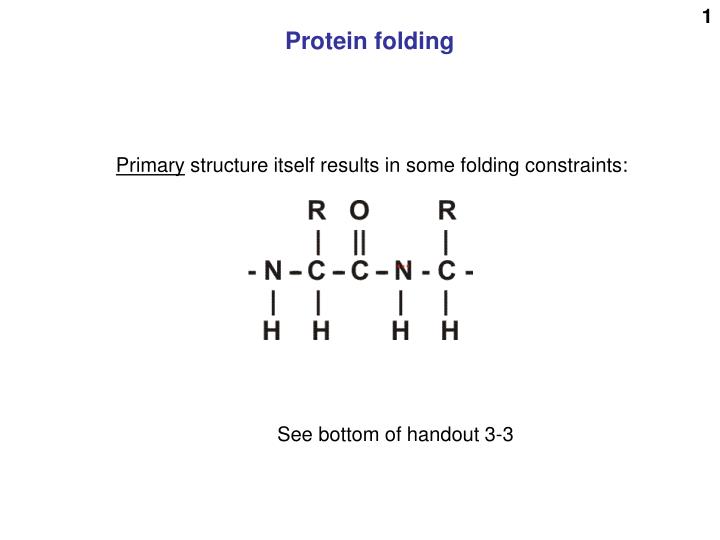
Protein folding. Primary structure itself results in some folding constraints:. See bottom of handout 3-3. And these 4 atoms are in one plane (N central). These 4 red atoms are in one plane (C of C=O central). so 6 atoms in one plane. There’s still plenty of flexibility.

E N D
Protein folding Primary structure itself results in some folding constraints: See bottom of handout 3-3
And these 4 atoms are in one plane (N central) These 4 red atoms are in one plane (C of C=O central) so 6 atoms in one plane
Secondary structure: the alpha helix Amino acids shown simplified, without side chains and H’s. H Almost every N-H and C=O group can participate
Alpha helix depictions C = grays N = blue O = red Poly alanine Side chains = -CH3 (lighter gray) H’s not shown
H-bond AA residue beta pleated sheet Secondary structure:
Beta sheet (i.e., beta pleated sheet) antiparallel antiparallel parallel
Beta-sheets Anti-parallel Parallel
secondary structure (my definition): structure produced by regular repeated interactions betweenatoms of the backbone.
Neither 3 alpha helices Tertiary structure: The overall 3-D structure of a polypeptide. This is a popular “ribbon” model of protein structure. Get familiar with it. The ribbons are stretches of single polypeptide chains. A single ribbon is NOT a sheet. A beta sheet These “ribbon” depictions do not show the side chains, only the backbone
Tertiary structure (overall 3-D) ionic hydrophobic H-bond cys Ion - dipoleinteraction covalent Van der Waals Examples of bonds determining 3D structure Exist in loop regions and in regions of secondary structure
Disulfide bond (covalent, strong) ½ O2 Disulfide bond formation Sulfhydryl group R-CH2-SH HS-CH2-R R-CH2-S-S-CH2-R + HOH + cystine cysteine cysteine Two sulfhydryls have been oxidized (lost H’s) Oxygen has been reduced (gained H’s). Oxygen was the oxidizing agent (acceptor of the H’s). An oxidation-reduction reaction: Cysteines are getting oxidized (losing H atoms, with electron; NOT losing a proton, not like acids.) Oxygen is getting reduced, gaining H-atoms and electrons Actually it’s the loss and gain of the electrons that constitutes oxidation and reduction, respectively. No catalyst is usually needed here.
Overall 3-D structure of a polypeptide is tertiary structure Stays intact in the jacuzzi at 37 deg C Usually does not require the strong covalent disulfide bond to maintain its 3-D structure [Tuber mode]l
Protein structures are depicted in a variety of ways Backbone only Ribbon Small molecule bound Drawing attention to a few side groups Continuous lines, ribbons= backbone (not sheets) Space-filling Space-filing, with surface charge blue = + red = -
Two different proteins with almost the same 3-D structure ! Handout 4-2
The usual weak bonds 4o, QUATERNARY STRUCTURE Monomeric protein (no quaternary structure) Dimeric protein (a homodimer) Dimeric protein (a heterodimer) Also called: multimeric proteins A heterotetramer A heteropolymeric protein (large one)
$ $ $ $ Hemoglobin One protein Four polypeptide chains, 2 identical alphas and 2 identical betas Four “subunits” Molecular weight 16,000 Subunit molecular weight 16,000 Subunit molecular weight Protein molecular weight 64,000 $ 64,000, even though the 4 chains are not covalently bonded to each other
The 4 weak bond types Tetramer Two heavy chains (H), Two light chains (L) Interchain disulfide bonds
glu val val glu glu val val glu Sickle cell disease Normal Sickle cell
Some small molecules can be bound tightly to a protein. Such associated small molecule are called “prosthetic groups”. Some are even covalently bound to the protein. Pyridoxal phosphate AAside chain Enzyme = Vitamin B6
Most prosthetic groups are bound tightly via weak bonds. Tetrahydrofolic acid ~ vitamin B9 Riboflavin~ vitamin B2 Heme
Membrane proteins Hydrophobic side chains on the protein exterior for the portion in contact with the interior of the phospholipid bilayer. Anions are negatively charged. Cations are positively charged
Small molecules bind with great specificity to pockets on protein surfaces Too far
Ligand Protein
Ligand binding can be equisitely specific: the estrogen reeptor binds estrogen but not testosterone. Testosterone Estrogen
Protein separation methods Ultracentrifugation Mixture of proteins
Causing sedimentation: centrifugal force = m(omega)2r m = mass omega = angular velocity r = distance from the center of rotation Opposing sedimentation = friction = foV. fo = frictional coefficient(shape) V = velocity Constant velocity is soon reached; then, no tnet force So: centrifugal force = frictional force (balanced each other out) And so: m(omega)2r = foV And: V = m(omega)2r/fo, Or: V = [(omega)2r] x [m / fo] V proportional to mass (MW) V inversely proportional to fo V inversely proportional to “non-sphericity” (spherical shape moves fastest)
Sample loaded here (“native”) Glass plates Winner: Small, +++ High positive charge + + + Loser: Large, + low positive charge +++ +++ +++ poly- acrylamide fibers + Intermediate: Large, +++ high positive charge + + Intermediate: Small, + Low positive charge +++ +++ +++ Molecules shown after several hours of electrophoresis
Upper resevoir Cut out for contactof buffer with gel
Cut out of glass platefor contact of buffer with gel Clamped glass sandwich Electrodeconnection(~ 150 V) Reservoir for buffer
Power supply Happy post-doc Tracking dyes
sodium dodecyl sulfate, SDS (or SLS): CH3-(CH2)11- SO4-- CH3-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-SO4-- SDS PAGE = SDS polyacrylamide gel electrophoresis SDS All the polypeptides are denatured and behave as random coils All the polypeptides have the same charge per unit length All are subject to the same electromotive force in the electric field Separation based on the sieving effect of the polyacrylamide gel Separation is by molecular weight only SDS does not break covalent bonds (i.e., disulfides) (but can treat with mercaptoethanol for that) (and perhaps boil for a bit for good measure)
Disulfides between 2 cysteines can be cleaved in the laboratory by reduction, i.e., adding 2 Hs (with their electrons) back across the disulfide bond. One adds a reducing agent: mercaptoethanol (HO-CH2-CH2-SH). In the presence of this reagent, one gets exchange among the disulfides and the sulfhydryls: Protein-CH2-S-S-CH2-Protein + 2 HO-CH2CH2-SH ---> Protein-CH2-SH + HS-CH2-Protein + HO-CH2CH2-S-S-CH2CH2-OH The protein's disulfide gets reduced (and the S-S bond cleaved), while the mercaptoethanol gets oxidized, losing electrons and protons and itself forming a disulfide bond.
e.g., “p53” Molecular weight markers (proteins of known molecular weight) P.A.G.E. 12 18 48 80 110 130 160 140
Molecular sieve chromatography (= gel filtration, Sephadex chromatography) Sephadex bead
Molecular sieve chromatography Sephadex bead
Molecular sieve chromatography Sephadex bead
Molecular sieve chromatography Sephadex bead
Molecular sieve chromatography Sephadex bead
Plain Fancy 4oC (cold room)
















