Download

1 / 104
1.06k likes | 1.45k Views
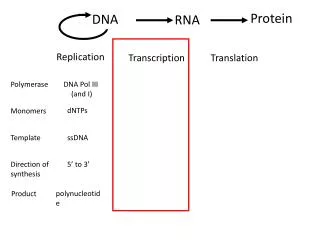
第七章 RNA 加工. 在细胞内由 RNA 聚合酶合成的初级转录产物( primary transcript )往往需要经过一系列的加工,才能转变为成熟的 RNA 分子。原核生物的 mRNA 一经转录通常立即进行翻译,一般不进行转录后加工。但是稳定的 RNA ( tRNA 和 rRNA )都要经过一系列的加工才能成为有活性的分子。真核生物由于存在细胞核结构,转录与翻译在时间上和空间上被分隔开,其 RNA 前体的加工过程极为复杂。.

E N D
第七章 RNA加工 在细胞内由RNA聚合酶合成的初级转录产物(primary transcript)往往需要经过一系列的加工,才能转变为成熟的RNA分子。原核生物的mRNA一经转录通常立即进行翻译,一般不进行转录后加工。但是稳定的RNA(tRNA和rRNA)都要经过一系列的加工才能成为有活性的分子。真核生物由于存在细胞核结构,转录与翻译在时间上和空间上被分隔开,其RNA前体的加工过程极为复杂。
RNA加工包括5’端和3’端的成熟、核苷的修饰、拼接和编辑等过程。另外,真核生物同一种前体mRNA通过外显子的不同连接方式可以形成两种或两种以上的mRNA。因此,对真核生物来讲,RNA的加工尤为重要。RNA加工包括5’端和3’端的成熟、核苷的修饰、拼接和编辑等过程。另外,真核生物同一种前体mRNA通过外显子的不同连接方式可以形成两种或两种以上的mRNA。因此,对真核生物来讲,RNA的加工尤为重要。
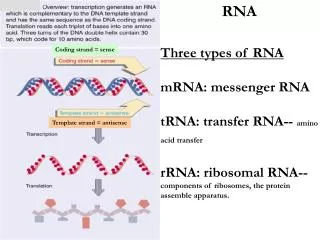
第一节 真核生物前体mRNA的加工 真核生物编码蛋白质的基因的初级转录产物称为前体mRNA,经过5’端加帽、3’端剪切及加多聚A尾、剪接和甲基化产生出成熟的mRNA分子。
一、5’端加帽 加帽是一个多步骤加工过程。当前体mRNA从RNA聚合酶II中伸出其5’端时即开始加帽反应。第一步是RNA 5’端的γ-磷酸基团由RNA三磷酸酯酶(triphosphatase)去除。然后在鸟苷酰转移酶的作用下,RNA末端核苷酸的β-磷酸基团亲核进攻和GTP的α-磷酸基团,产生5’-5’对接的磷酸二酯键,同时释放出焦磷酸。最后一步反应是在鸟嘌呤甲基转移酶的作用下将一个甲基基团加到鸟嘌呤环的第7位N原子上,使鸟嘌呤转变成7-甲基鸟嘌呤。
7-甲基鸟嘌呤结构称为0型帽子(type 0 cap),是酵母中最常见的形式。在高等真核生物中5’端还会发生更多的修饰。在转录产物的第一个核苷酸核糖的2’-OH被甲基化,形成I型帽子。脊椎动物转录产物的第二个核苷酸核糖的2’-OH也被甲基化,则形成II型帽子。
2、加帽的功能 mRNA 5’加帽的功能主要表现在4个方面: 1.阻止mRNA的降解:细胞内存在许多RNA酶,它们可从5’端可攻击游离的RNA分子。当mRNA的5’端加上m7GpppG帽子后,可阻止RNase的切割,延长mRNA的半衰期。 (2)提高翻译效率 真核生物mRNA必需通过5’帽结合蛋白才能接触核糖体,起始翻译。缺少加帽的mRNA由于不能被5’帽结合蛋白识别,其翻译效率比加帽的mRNA低20倍。
(3)作为进出细胞核的识别标记 凡由RNA聚合酶II转录的RNA均在5’端加帽,包括snRNA,这是RNA分子进出细胞核的识别标记。U6 snRNA 由RNA聚合酶III转录,其5’端保留3个磷酸基团,无帽子结构,因而不能输出细胞核。 (4)提高mRNA的剪接效率 5’帽结合蛋白涉及第一个内含子剪接复合物的形成,直接影响mRNA的剪接效率。
二、3’端加尾与转录的终止 加尾信号 真核细胞中几乎所有的成熟mRNA的3’末端都有一串腺苷酸构成的尾巴。这些腺苷酸并非由DNA编码,而是在mRNA 3’端成熟的过程中由Poly (A)聚合酶添加到mRNA分子上的。前体mRNA 3’端的加工需要信号序列的指导。靠近mRNA 3’端有一加尾信号5’-AAUAAA-3’。加尾信号下游10~30 nt处有一5’-CA-3’二联体。二联体的后面有一段富含GU的序列。
cleavage site 5’-cap Coding sequence AAUAAA GU-rich 10 – 30 nts 20 – 40 nts Poly-adenylation CA Endonuclease AAUAAA CA ATP PPi Polyadenylate polymerase RNA+ nATP = RNA-(AMP)n + nPPi AAUAAA CA AAA(A)n 80 – 250 A’s
2、加尾反应 mRNA 3’端加尾实际上涉及到切割和加尾两个性质不同的反应,并且有多种蛋白质的参与。切割与多聚腺苷酰化特异性因子(cleavage and polyadenylation specificity factor, CPSF)与加尾信号5’-AAUAAA-3’结合。切割激发因子(cleavage stimulation factor, CstF)特异性地附着于GU丰富区。
CPSF和CstF结合位置处于切割与多聚腺苷酸化位点两侧,它们为切割因子、Poly A聚合酶以及Poly A结合蛋白的组装提供了平台。一旦聚腺苷酸化复合体组装完成,由切割因子I和切割因子II构成的内切核酸酶对RNA进行切割, 切割位点就位于5’-CA-3’二核苷酸的后面。Poly A聚合酶(Poly A polymerase)在新生的3’末端添加多聚A尾巴。Poly A结合蛋白与多聚A尾巴结合。
3’-End Formation: RNA Processing, Not Transcription Termination per se
1.多聚腺苷酰化特异性因子(cleavage and polyadenylation specificity factor, CPSF) 2.切割激发因子(cleavage stimulation factor, CstF) 3.切割因子I和切割因子II 4. Poly A聚合酶(Poly A polymerase)
3、RNAP II的终止反应 加尾信号同时指导转录的终止反应。当加尾信号被传送到延伸复合体时,EC的构象会发生相应的变化,导致具有抗转录终止作用的延伸因子(Paf1C, PC4)脱离EC。同时,与转录终止反应有关蛋白质因子,例如Xrn2,与EC结合。Xrn2是一种5'-3' 外切核酸酶,一旦发生切割反应,Xrn2就结合到下游RNA分子的5' 末端。Xrn2一边降解RNA,一边追赶Pol II,这一过程可能需要RNA/DNA解旋酶SETX的协助。当Xrn2赶上Pol II,会介导Pol II与模板脱离终止转录过程。与Pol I和Pol III一样,在转录终止之前Pol II也会在加尾信号之后出现停顿。
三、剪接 真核生物的基因常常是不连续的,即编码区被非编码区打断,具有这种结构的基因称为断裂基因(split gene)。其中,编码区称为外显子,非编码区称为内含子。外显子和内含子都能被转录,形成前体mRNA。在细胞核中,前体mRNA要被加工为成熟的mRNA。加工时,内含子被切除,外显子被连接在一起,此过程称为剪接。
1. 剪接信号及两次转酯反应 比较cDNA和基因组DNA序列可以确定前体mRNA分子中外显子和内含子的边界。在比较大量的真核生物的内含子序列后,发现大多数内含子的5’端的两个核苷酸是GU,3’端的两个核苷酸为AG,这类内含子也被称为“GU-AG”内含子,它们中的所有成员都以相同的机制进行剪接。
剪接信号:5’剪接位点;3’剪接位点;嘧啶丰富区;分支位点剪接信号:5’剪接位点;3’剪接位点;嘧啶丰富区;分支位点
分支位点A与内含子的第一个核苷酸(G)形成2’, 5’-磷酸二酯键,内含子自身成环,形成套索结构。
3. 剪接过程 哺乳动物的细胞核中有6种富含U的小RNA,被命名为U1、U2、U3、U4、U5和U6 snRNA,这些RNA的长度在107至201 nt之间,分别与蛋白质结合形成核内小核糖核蛋白(small nuclear ribonuleoprotein, snRNP)。这些snRNP与其他蛋白质因子一起按严格的程序组装成剪接体(spliceosome)完成两步剪接反应。
4、外显子剪接增强子 在内含子的剪接过程中,剪接装置必须识别正确的剪接位点,以保证外显子在剪接的过程中不被丢失,同时荫蔽的剪接位点要被忽略。所谓隐蔽剪接位点(cryptic splice site )是指与真正的剪接位点相似的序列。已经知道一类被称为SR蛋白(SR protein)的剪接因子在剪接位点的选择中发挥重要作用。
SR蛋白因它们的C端结构域有一个富含Ser(S)和Arg(R)的区域而得名。SR蛋白结合到外显子中外显子剪接增强子(exonic splicing enhancer, ESE)。与ESE位点结合的SR蛋白将U2AF蛋白引导到3’剪接位点,并将U1 snRNP引导到5’剪接位点,这是剪接过程的一个关键步骤,该步骤决定着哪些位点将被连接起来。
5、II类内含子的自我剪接 II类内含子存在于真菌和植物的细胞器基因组中,少数存在于原核生物的基因组中。自我剪接的意识是前体RNA中的内含子自身折叠成一种特殊的构象,然后催化自身的释放的过程。 II类内含子的剪接机制与核基因内含子的剪接具有一定的相似性。剪接是由内含子的一个保守的腺苷酸发动的。该腺苷酸的2’-OH作为亲核基团攻击内含子5’端的磷酸二酯键,使其断裂,内含子形成套索结构。
上游内含子的3’-OH作为亲核基团攻击内含子3’剪接位点,上游外显子和下游外显子连接起来,释放出内含子。在试管中,在没有任何蛋白质或其他RNA分子存在的情况下,上述两种转酯反应能够由II类内含子独立完成。上游内含子的3’-OH作为亲核基团攻击内含子3’剪接位点,上游外显子和下游外显子连接起来,释放出内含子。在试管中,在没有任何蛋白质或其他RNA分子存在的情况下,上述两种转酯反应能够由II类内含子独立完成。
5. 可变剪接(alternative splicing) 真核生物基因经转录后,其初级转录产物在剪接过程中呈现出不同的方式。有些基因的初级转录产物经剪接只产生一种成熟的mRNA。还有一些基因的初级转录产物经过不同的剪接途径,产生多种成熟的mRNA,继而产生功能不同的蛋白质。一般把一个mRNA前体经过不同方式的剪接产生相关但不同的成熟的mRNA的过程称为可变剪接(alternative splicing)。
哺乳动物肌钙蛋白T基因初级转录产物具有5个外显子。该pre-mRNA可以通过两种途径进行剪接,产生两种不同的mRNA。一种mRNA含有外显子1、2、3和外显子5,编码α肌钙蛋白T,另一种mRNA含有外显子1、2、4和外显子5,编码β肌钙蛋白T。哺乳动物肌钙蛋白T基因初级转录产物具有5个外显子。该pre-mRNA可以通过两种途径进行剪接,产生两种不同的mRNA。一种mRNA含有外显子1、2、3和外显子5,编码α肌钙蛋白T,另一种mRNA含有外显子1、2、4和外显子5,编码β肌钙蛋白T。
在较高等的真核生物中,可变剪接非常普遍。当果蝇的基因组序列草图完成以后,人们发现果蝇的基因数目比线虫的要少,尽管果蝇的解剖学结构比线虫复杂的多。生物的表型最终是由蛋白质决定的,有机体组织结构的复杂性反映着蛋白质组的多样性。果蝇基因组的基因数目与它的蛋白质组的蛋白质数目之间不一致的现象可用mRNA的可变剪接来解释。mRNA可变剪接极大地丰富了蛋白质组的多样性。根据人类蛋白质组的大小,预期人类的基因数目是80 000-100 000个,然而人类基因组草图顺序只给出了大约35 000个基因。现在知道至少有35%的人类基因进行可变剪接。
同一RNA分子的内含子被除去,外显子连接在一起的剪接称为顺式剪接(cis-splicing)。剪接不但能够发生在同一RNA分子上,也可以发生在不同的RNA分子间。不同RNA分子上的两个外显子剪接在一起的过程称为反式剪接(tran-splicing)。同一RNA分子的内含子被除去,外显子连接在一起的剪接称为顺式剪接(cis-splicing)。剪接不但能够发生在同一RNA分子上,也可以发生在不同的RNA分子间。不同RNA分子上的两个外显子剪接在一起的过程称为反式剪接(tran-splicing)。
1982年Piet Borst对锥虫衣被蛋白(surface coat protein)基因及其编码的mRNA的5’末端进行了测序,发现二者并不匹配:mRNA具有一段在衣被蛋白基因中不存在的35 nt序列。事实上,锥虫的所有mRNA都具有相同的35 nt的前导序列。该序列被称为剪接前导RNA(splicing leader)。SL由一个独立的基因编码,该基因在锥虫的基因组中大约重复200次,它的编码产物由两部分组成:SL以及其后的100 nt序列。
锥虫的基因排列成巨大的多顺反子转录单位,转录后形成多顺反子RNA。通过反式剪接,单一的成熟mRNA从多顺反子RNA中释放出来。反式剪接反应发生在SL基因的转录产物和多顺反子RNA之间。在SL基因转录产物中SL序列和其后的100 nt序列之间为5’剪接位点,多顺反子RNA每一编码区和其上游间隔序列的交界处为3’剪接位点。发生剪接反应时,5’剪接位点GU和3’剪接位点AG上游的分支位点之间形成2’ -5’连接的磷酸二酯键。接着发生第二次转酯反应,SL序列连接在基因编码区的5’端,同时释放出Y型分子。
四、RNA编辑(RNA editing) RNA编辑(RNA editing)是RNA加工的一种形式,它通过碱基替换、插入或删除使原始转录产物核苷酸序列被更改。