Download

1 / 39
390 likes | 422 Views
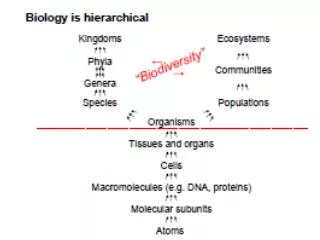
Biodiversity Ricklefs’ definition: Biodiversity is “a measure of the variety of organisms within a local area or region, often including genetic variation, taxonomic uniqueness, and endemism. Ricklefs is slightly more precise when he defines diversity as an ecological measure… Diversity is:

E N D
Biodiversity Ricklefs’ definition: Biodiversity is “a measure of the variety of organisms within a local area or region, often including genetic variation, taxonomic uniqueness, and endemism. Ricklefs is slightly more precise when he defines diversity as an ecological measure… Diversity is: a) the number of taxa in a local area or region or b) A measure of the variety of taxa in a community that takes into account the relative abundance of each one.
Definition a is usually called species richness. As a count of the number of species, it does not take into account relative abundance. It would claim the diversity of a community with 100 of species A and 1 each of species B,C,D, and E was 5, the same as one with 20 individuals of each species. Depending on the details of the measure used, definition b may be called a dominance index or, if information theory is used to generate the index, it is called Shannon’s or the information theory diversity index. The dominance index for a community is calculated as:
The Shannon information theory diversity index is calculated • as: • Since H is an index related to logs, two ‘corrections’ have • been suggested for it: • Instead of using H, compare it as a ratio to the maximum • possible diversity for the community (if all species were • equally abundant, called H’). This ratio is called evenness. • b) Make the value more proportional to the number of species • by using the H as the exponent of an expression that doesn’t • have an accepted name. The reported diversity is then 2H. Note that your text (pg. 417) suggests using logs base e. The result is comparatively identical, but isn’t an information theory measure.
Here’s a sample calculation of the two diversity indices for a community containing 5 species: Species richness = 5 Relative abundances: Species # individuals pi pi2 A 15 .123 .015 B 20 .164 .027 C 75 .615 .378 D 4 .033 .001 E __8___ .066 .004 Total 122 0.425 D = 1/0.425 = 2.352 H = 1.142 (using ln rather than log base 2)
Obviously, a more diverse community has more species and no single (or a few) species predominant in the community. But what determines how many species we are likely to find, and what determines how many species there are in the world? The answer to the second question is fairly straightforward… There are about 2 million species that have been officially named. Some groups are well known, others have only a small fraction named. You can probably guess which are well known:
Birds – there are about 9000 species. Half had been named by the middle of the 19th century. Today a few species are named each year. Mammals – there are about 4000 species, and less than 10 are added each year (almost all small rodents). Are they the most diverse animal fauna? NO!
Globally, the most accepted estimates of total taxonomic diversity expect that if we could find and name them all, we’d have somewhere between 10 and 30 million species. The majority of the total are insects, and among insects the largest number are beetles. J.B.S. Haldane wrote that “The creator has an inordinate fondness for beetles.” About 20% of all species are beetles, and we find new species frequently and in large numbers… Terry Erwin has collected beetles from trees in tropical forests for many years. In one series of studies he used insecticidal fogging to collect insects in the Tambopata Reserve in Panama. The results are staggering.
The species overlap between two plots only 50m apart in his study area was only 8.7% (i.e. 91.3% of the beetles were different over this short distance). There were 163 species of beetles unique to one species of tree, Luehea seemannii, in its canopy. Since beetles make up about 40% of all insects, Erwin estimated that this tree canopy harbored ~400 unique species of insects (160 beetles and 240 other insects). What about the other parts of the tree – trunk and roots? We can only guess how many others may live there. Estimates (E.O. Wilson 1992) suggest about twice as many species in the canopy as on and in the ground. So, there are maybe 600 species of insect unique to this tree species. There are an estimated 50,000 tropical tree species, so…
There may be 30,000,000 tropical insect species. What about the rest of the world? Tropical rainforest is the most diverse biome on earth. Many fewer species would be added by expanding the range of habitats included. There are also errors of unknown size in each step of the estimation process. So, with no better information to make an estimate of total global diversity more precise or more accurate, the loose estimate of from 10 – 30 million is a useful start.
While it is generally accepted that tropical rainforest is the most diverse biome on earth, there are also indications of a large scale pattern in diversity globally. In general, the diversity of species declines with latitude, i.e. as you move from the tropics toward the poles. Your text uses the number of ant species to show the pattern… Amazon rain forest
Many taxa have been surveyed in North America. Mammals also fit the latitudinal pattern, so do lizards. In each case equal areas are compared, so it isn’t just that the range is larger at lower latitude… Mammals – lizards
Ecologists have generated a number of different (but in many cases not completely independent) hypotheses to explain the latitudinal gradient in diversity. We’ll consider the hypothesis in turn. They are: • Evolutionary time 8. Competition • Ecological time 9. Disturbance • Climatic stability 10. Predation • Climatic predictability • Spatial heterogeneity • Productivity • Stability of primary production
These mechanisms are likely working at different spatial scales. At the geographical or regional scales: evolutionary time ecological time climatic stability or predictability hypotheses invoking increased resource partitioning spatial heterogeneity competition productivity and stability of productivity At the local scale: disturbance predation
Evolutionary time – based on the assumption that diversity increases with the age of a community. In the northern hemisphere, relatively recent glaciations mean that temperate and polar communities are very recent compared to tropical communities. This is argued to explain their relative paucity in species. However, we now know that, though tropical rainforest persisted through the 4 cycles of Pleistocene glaciation, the community was not untouched. Rainfall patterns changed during glacial epochs, and continuous tropical rainforest was fragmented during glaciation (in the Amazon Basin into at least 7 separate continuing forest areas; the areas between were probably savannah.). So, there are old and relatively recent communities in tropical rainforest.
Here’s the simple view of what the evolutionary time hypothesis suggests:
Ecological time – this hypothesis suggests that the time since glaciation has been too short for dispersal of species to occupy their full range of habitats (and latitudes). When glaciers receded, there was newly open habitat. How rapidly can species migrate/disperse to occupy suitable places? Dispersal capacity for most species suggest that this is at most a minor factor.
Climatic stability – Here reference is made to the amplitude of the seasonal cycle of climate. If there is little seasonality, species can live there with little need for broad tolerance capability (a broad niche with respect to physical climatic variables). However, if there is high amplitude seasonal variation, either broad tolerances or other adaptations are necessary. If niches are broad, how many species can be crowded in? Fewer than if niches are narrower. Other adaptations? These can include hibernation, diapause (loosely, insect hibernation), dormancy (plants), and, possibly, the evolution of migration.
Along the same gradient (physical conditions or some resource) you can pack more species with narrow niches… 3 spp. Species Abundance breadth Resource or climatic gradient 6 spp. Species Abundance Resource or climatic gradient
Climatic predictability – If a climate has high amplitude cycles, but those cycles occur predictably each year, then adaptations can be selected to manage the problem. Most (if not all) of the “other” adaptations to climatic stability may well fit better here, as adaptations to predictable cycles. Species may time important phases of the annual life cycle to fit environmental conditions. For example, in the desert of the American southwest, there are two periods when rainfall is more likely…
Here’s a comparison between rainfall at Omaha, Nebraska and Phoenix, Arizona. Note the difference in the scales along the y- axes. In the southwest desert there are two groups of plants. One’s seeds germinate in the cool-wet (winter annuals), and the other’s germinate in warm-wet, and flower after summer flash floods.
Spatial heterogeneity – there is a strong correlation between the structural complexity of a habitat and the number of species it holds. Text figure 23.5 shows this for birds from deciduous forest in eastern North America… Just to show you this wasn’t a carefully chosen habitat, the same relationship is found for birds over a wide range of habitats in both North America and Australia…
If there are more possible specializations in habitat conditions (indicated by such measures as foliage height diversity), then more species may be able to co-occur by specialization on a particular part of the ‘gradient’. sp. 1 sp. 2 sp. 3 Fewer resources or possible habitat specializations, so fewer species Species Abundance Resource More resources or…, more species sp. 1 sp. 2 sp. 3 sp. 4 sp. 5 sp. 6 Species Abundance Resource
Productivity – More productivity means more food available, and the potential for greater specialization. Thus, everything shown on those last graphs applies here, as well. One way of seeing this is to look at how a graph of resources available can be subdivided into equal areas, and how many such areas pack in when there are more resources available.
Your text (fig.23.3) shows that productivity and habitat complexity interact to some degree. High productivity without a means to specialize (complexity) will simply lead to larger population sizes rather than increased diversity.
Sometimes productivity is estimated by using potential evapotranspiration. This is a measure that combines evaporation from the soil surface and transpiration. It indicates energy input into a system. There is a pattern relating PET to species richness in a number of major taxonomic groups. Ricklefs shows the relationships for North America.
Stability of primary production – Species can only successfully partition resources finely if the resources do not vary too substantially in abundance over time. Some call this hypothesis one of temporal heterogeneity to parallel the spatial heterogeneity hypothesis, but clearly with an opposite result on diversity. In the attempt to avoid predation on fruits and/or seeds, some plants attempt to become ‘unpredictable’ in providing resources by adopting a strategy of masting, producing a heavy crop one year, but very little the next. That unpredictability should reduce the diversity (and number) of granivores or frugivores feeding from the plant.
Competition – this hypothesis has a lot of overlap with others that invoke small niche breadth as being key to high diversity. For example, it is believed that tropical forest species are highly K-selected. Competition is fierce, the populations are near their equilibria in the face of both intra- and interspecific competition. There is strong selective pressure to reduce niche breath to a region of niche space where a species is superior. The resulting narrow niches allow more species to be packed into the same resource space. Just the opposite – broader niches, lower diversity – would be expected where r-selection predominates, i.e. harsh, high latitude habitats.
Disturbance (and the intermediate disturbance hypothesis) – Disturbances reduce the density of individuals, and thus the intensity of competition. You can consider this to be the inverse of the competition hypothesis in many senses. The intermediate disturbance hypothesis says that when disturbances are rare, the community becomes filled with the K-strategists. When disturbances are frequent, those capable of rapid growth to recover following the disturbance should predominate, i.e. r-strategists. At an intermediate frequency and intensity of disturbance, members of both groups may be present, and thus diversity may be maximized. Here’s a figure abstracting this idea…
When disturbances occur frequently, populations can never grow to an abundance where resources become limiting. Thus, competition among the species cannot be important. However, species that are slow to mature and reproduce cannot tolerate frequent disturbance, i.e. K-strategists. The species that can tolerate frequent disturbance are those that mature and reproduce rapidly. Those species, r-strategists, also tend to have high rates of dispersal, and are therefore likely to ‘find’ disturbances soon after they occur.
If disturbances occur rarely, then populations do reach a size where resource limitation is important. Thus, competition among the species occurs, and dominant competitors can exclude poorer competitors from the community. Species diversity is lower due to the loss of these inferior competitors. But, at an intermediate frequency of disturbance (and each place I’ve talked about frequency you could substitute intensity) both r- and K-types may persist together, and diversity should be higher. There is evidence both supporting and not supporting the intermediate disturbance hypothesis…
First, evidence that doesn’t work: the effect of burning on plant species diversity on Konza Prairie in Kansas. Here, species diversity declines with increasing frequency of burning, but there is no intermediate peak…
But here is evidence that supports the hypothesis. It was initially constructed by Joe Connell for the intertidal. Here it was applied by Wayne Sousa to colonization of shoreline boulders by green algae. Each line is a species. Ulva, the line that starts first, is a pioneer. Note that it has disappeared by the end. Both the species that arrive late and Ulva are present at the middle of the sequence…
Sousa also knew that wave action (the main disturbance force), would be most likely to roll small boulders (frequent disturbance), least likely to roll large ones, and roll medium sizes and intermediate number of times. What was the diversity on the different boulder classes?
Predation – here the hypothesis parallels what was described for keystone predators. The intensity of competition among prey species is reduced, thus permitting a greater number of species to persist. This is especially true when the predators follow a type III functional response. If predation is prey frequency dependent, then prey diversity can increase.
When we talk about diversity, we have to careful to identify the scale we are talking about. Within a single community, there is a diversity of species present. At the local level, we call this α diversity. There is a second, larger level in which we identify the variation in the species lists of communities in different habitats within a region. This is β diversity. Finally, there is a third level that considers the total species diversity over the whole region. This level is called γ diversity. γ diversity = average α diversity x β diversity
This relationship can be viewed in a slightly different way. γ diversity indicates the entire species pool available in a region. Not all species from this pool are present in each community. Instead, there is a process called species sorting that separates the total pool into groups of species that can successfully coexist in single communities. One of the best (and few) studies to investigate sorting was work done by Paul Keddy of the University of Ottawa. He planted seeds of 20 different wetland species (his regional pool) into 120 different wetland areas, among which physical and chemical conditions differed. He then followed what happened for 5 years…
One species failed to germinate in any of the 120 sites. 5 others didn’t persist in any community for the full 5 years. That left 14 species to be sorted by the differences in physical and chemical conditions of the individual plots. Ricklefs plotted the results in terms of the average number of species in individual plots, separated into those from plots with fertile soil and those from infertile soil. fertile soil