Download
1 / 37
370 likes | 609 Views
Chemical Biology I TIGP0101-00 Enzyme Kinetics B807 TEL:27898662 Institute of Chemistry

E N D
Chemical Biology I TIGP0101-00 Enzyme Kinetics B807 TEL:27898662 Institute of Chemistry Academia Sinica Fall, 2003 • Mechanisms: • A P • Kinetics: steady state/pre steady state • Spectroscopic: structure(or active site) NMR/EPR (fluorescence) • X-ray: structure determination at active site • Binding studies: thermodynamic understanding inhibitors • transition states. • sequence analysis, genomics, • genetic manipulation • DNA protein sequence mutants
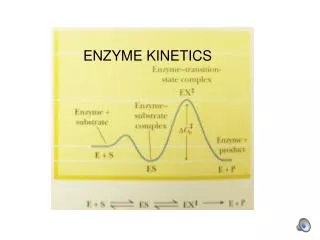
Enzymes: A rate P T.S. overcome energy barrier: (1). lower barrier stabilize T.S. (2). destabilize ground state or enzymes substrates (A) A P • rate acceleration: how fast? • specificity: how selective?
destabilize ground state or enzymes substrates (A) lower barrier stabilize T.S.
Rate acceleration: non-enzyme very slow How fast of reaction rate enzyme will facilitate? Catalytic Power *turnover number: the number of substrate molecules converted into product per enzyme molecule per unit time when the enzyme is saturated with substrate. Vmax moles/min Et moles enz. min-1 1 106, s-1 usually average’s rate, 6 s-1 EX1: Urease EX2: Catalase EX3: Carbonic anhydrase
Specificity: Enzymes’ active envolved to do some specific things. EX1: Hexokinase phosphorylation: EX2: Alcohol dehydrogenase:
Enzyme commission Systematic Name number • Oxidoreductases(oxidation-reduction • reactions) • Transferases(transfer of functional groups) • Hydrolases(hydrolysis reaction) • Lyases(addition to double bonds) • Isomerases(isomerization reactions) • Ligases(formation of bonds with ATP cleavage) Example: EC 1.1.1.1 alcohol dehydrogenase EC 2.1.1.1 nicotinamide N-methyltransferase EC 3.3.1.21 -glucosidase EC 4.1.1.1 pyruvate decarboxylase EC 5.3.1.1 triose-phosphate isomerase EC 6.5.1.3 RNA ligase
zero-order on [A] K1A K3 E EA P + E K2 EP d[EA] dt d[E] dt = 0 = 0 Steady state: A P rate x x x x x x mixed order x x x first order on [A] x [A] Steady-state assumption: 1925, G. E. Briggs and James B. S. Haldance assuming the concentration of the enzyme-substrate complex (EA) quickly reaches a constant value in such a dynamic system. That is, EA is formed as rapidly from E + A as it disappears by its two possible fates: dissociation to regenerate E + A, and reaction to form E + P.
A + B P + Q Nomenclature: by Cleland substrates A, B, C, D,.....etc products P, Q, R, S,......etc inhibitors I, J, K,......etc enzyme complex E, F, G (stable complex) enzyme complex EA (unstable transitory complex) enzyme complex EAB EPQ (central complex) E : free enzyme F : covalent attachement enzyme complex
K1A K3 VmaxA E EA P + E K2 EP Ka + A 1 Ka + A Ka 1 = = + reciprocal VmaxA Vmax VmaxA rate x x x x x x x Steady state: Michaelis Menten equation = x x first order on [A] x [A]
K1A K3 E EA P + E K2 EP Ka 1 dP dt = k3 [EA] rate = Derivation of Rate Equations (Biochemistry, 1975, 14, 3320) 1/ slope k/V 1/V 1/A
K1A K3 E EA P + E K2 EP d[E] dt dP dt d[EA] dt d[E] dt d[EA] dt dP dt = [E]K1A K2[EA] K3[EA] = K3[EA] K2[EA] K1A[E] = k3 [EA] = k3 = k3 [EA] = 0 = 0 rate = rate = ET = E EA Steady state assumption: Solve for EA E = ET EA K1A(ET EA) K2(EA) K3(EA) = 0 K1AET K1A(EA) K2(EA) K3(EA) = 0 K1AET= EA(K1A K2 K3) EA = K1AET (K1A K2 K3) because K1AET (K1A K2 K3)
k3A = VmaxA = K2 K3 A Ka+ A K1 dP dt = k3 [EA] = k3 rate = K1AET (K1A K2 K3) divide by K1 k3K1A rate = (K1A K2 K3) ET • rate as A , k3 is predominate, k3 =Vmax • (2)rate as A 0, K1A 0 , rate k3K1A = (K2 K3) ET (initial rate) k3K1 V = because K3 = Vmax (K2 K3) K (K2 K3) K = K1
K1A K3 K5 Steady-state Rate Law for a One-substrate, One-product Reaction with Two Reversible Steps A P EP E EA P + E K2 K4 binding chemical dissociation Replacing every equilibrium rate constant by net rate constant: Net rate constant: E1 E2 E3 E1 K1 K3 K5 steady state each [E] depend on next net rate constant K magnitude if K3 large, [E2] if K3 small, [E2] 1 E1 K1 1 E1 Therefore, , = K1 Et 1 1 1 + + K1 K3 K5
Flux is constant at steady state: rate =E1(K1) =E2(K3) =E3(K5) at steady state ET velocity = E1(K1) = 1 1 1 + + K1 K3 K5 1 E1 K1 {because } = Et 1 1 1 + + K1 K3 K5 1 = n 1 Et Ki i <Homework> Go back to derive an equation for a one-substrate,one-product reaction with one reversible steps
1 Ka + A Ka 1 = = + VmaxA Vmax VmaxA Lineweaver-Burk double-reciprocal plot
K5 K6 Kinetic Mechanisms forward V1 ; reverse V2 Michaelis complexes Ka, Kb inhibition constants(thermodynamic) Kia, Kib (A). Sequential mechanism: All substrates bind before chemical events. 1. Order: Enzyme binds in different order with substrates. If the mechanism is ordered, the substrates will add to the enzyme as A first, B second, etc., and the first product to dissociate from the enzyme will be P, followed by Q etc. (a). Order sequential mechanism: NAD+-dependent dehydrogenases E EA EAB K9 K1A K3B K7 EQ E EAB EPQ E EA K2 K10P K4 K8P
Order sequential mechanism: A B P Q E EA (EAB EAP) EQ E *It may be impossible for B to bind until after A binds and promotes a conformational change in the enzyme that exposes the B binding site. (b). Theorell-Chance mechanism:steady state concentration of central complexs are low. A B P Q E EA EA E example: liver alcohol dehydrogenase.
2. Random: A enzyme catalyzing a random mechanism would possess two distinct sites, one for each substrate(or product), so that the reaction of one substrate with the enzyme may occur before or after the other. EA E EAB EB A B P Q E E EAB EPQ B A Q P (a). Ordinary random mechanism: if slowest step is one other than the interconversion of the central complex, EAB EPQ. (no enzyme is known to have this mech.) (b). Random-rapid equilibrium mechanism: If the slowest is central complex. example:yeast hexokinase, creatine kinase.
(B). Ping Pong mechanism: Chemistry occurs prior to binding of all substrates The addition of one substrate to the enzyme causes a reaction which results in the formation of one product and a new stable form of the enzyme which in turn reacts with the second substrates. examples: thioltransferase, phosphoglucomutase transaminase. A P B Q E (EA FP) F FB EQ E a new stable form of the enzyme
Kinetics of Enzyme-catalyzed Reactions Involving Two or more Vary Substrates A + B P + Q • Intersecting Pattern: indicates sequential combination of both substrates prior to release of a product. 1/ 1/ [B] [A] 1/A 1/B V1AB = KiaKb + KaB + KbA + AB
Kinetics of Enzyme-catalyzed Reactions Involving Two or more Vary Substrates A + B P + Q 2. Parallel Pattern: An irreversible step intervenes between the times of combination of the two substrates in the mechanism. 1/ 1/ [A] [B] 1/B 1/A VAB = KaB + KbA + AB
Kinetics of Enzyme-catalyzed Reactions Involving Two or more Vary Substrates A + B P + Q 3. Equilibrium Ordered Pattern: Since it corresponds to ordered addition of A and B, with addition of A at equilibrium, looks different when [A] and [B] are varied. [A] [B] 1/ 1/ 1/B 1/A VAB = KiaKb + KbA + AB • This pattern is most commonly seen with metal activators which • are not consumed during the reaction, but must be present to • permit substrate binding.
Slope and Intercept intercept---velocity at sat. substrate , observe intercept. A B Slope---rate at low substrate concentration *Sequential mech. A B P Q E EA (EAB EAP) EQ E intercept change enzyme different A EA B EAB E EA slope will change if change [B]
Slope and Intercept *Ping pong mech. A P B Q E (EA FP) F FB EQ E intercept change slope no slope effect by change [B]
Enzyme Inhibition product, dead-end substrate inhibited enzyme 1. Competitive inhibition (C): A competitive inhibitor is a substance that combines with free enzyme in a manner that prevents substrate binding. That’s, the inhibitor and the substrate are mutually exclusive, often because of true competition for the same site. 1/ [I] 1/A Slope change only Vmax is the same
Active site of enzyme Competitive inhibition (C): Substrate Inhibitor Substrate and inhibitor can bind to the active site Products Inhibitor prevents binding of substrate
Enzyme Inhibition 2. Uncompetitive inhibition (UC): A classical UC inhibitor is a compound that binds reversibly to the enzyme-substrate complex yielding an inactive ESI complex. The I does not bind to free enzyme. K1 K3 1/ E + A EA P + E K2 + I [I] KI 1/A EAI NO REACTION Intercept change Slope is the same
Enzyme Inhibition 3. Noncompetitive inhibition (NC): A classical NC inhibitor has no effect on substrate binding and vice versa, A and I bind reversibly, randomly and independently at different sites. 1/ [I] 1/A Slope change Intercept change
Active site Noncompetitive inhibition (NC): Binding of inhibitor distorts the enzyme Inhibitor site Substrate and inhibitor can bind simultaneously In the absence of inhibitor, products are formed The presence of the inhibitor slows the rate of product formation
VmaxA VmaxA VmaxA Km + A Km+ ’A Km + A = = = Km Km ’ ’ 1 1 1 1 Km Km 1 1 = = = = + + + + VmaxA VmaxA VmaxA VmaxA Vmax Vmax Vmax Vmax VmaxA = Km+ ’A Effects of Inhibitors on Michaelis-Menten Reactions Lineweaver-Burk Equation Effect of Inhibitor Michaelis-Menten Equation Type of Inhibition None None Increase Km Competitive Decrease Km and Vmax Uncompetitive Decrease Vmax; may increase or decrease Km Noncompetitive = 1 + [I]/KI ’ = 1 + [I]/K'I
Intercept Idea: competitive pattern 1/ No inhibition by [I] if A 1/A 0 [I] I and A competiting for the same site (for the same enzyme) No intercept 1/A I and A bind to different enzyme intercept effect will become NC inhibition Exceptions:
Slope effect: K1A E EA K2 EA respect E lower E respect EA raised EA E I reversibly connected to either or show slope effect actual product inhibitors example: dead-end inhibitor
Catalysis • Covalent catalysis: rate acceleration from the formation of covalent bonds between enzyme and substrate. Enz-X: better attacking group and better leaving group example: ping-pong mechanism
2. Acid/base catalysis: (a) specific acid-base catalysis (b) general acid-base catalysis general acid-base catalysis This reaction accelerated by imidazole. Usually increasing concentration of product(imidazole) will decrease the rate. However, imidazole help to extract H+from water molecules in T.S.
3. Entropy: entropy loss in the formation of EA The rotational and translational entropies of the substrate have been lost already during formation of EA complex example: Strain/distortion
Transition state: Enzyme stablize T.S. to accelerate the reaction rate. Enzyme should bind tighter in T.S. than in substrate and product states. example: Proline racemase and Isocitrate lyase (Prof. Robert Abeles)