Download

1 / 148
1.48k likes | 1.78k Views
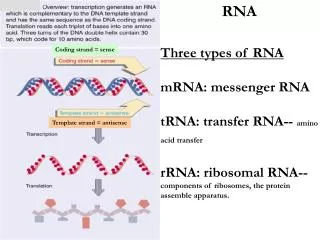
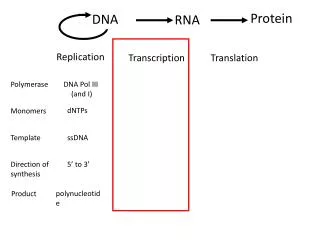
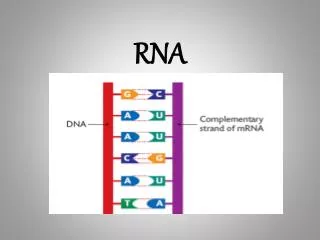
第十四章 RNA 的生物合成. DNA 转录 RNA 的复制。 RNA DNA RNA. RNA 的生物合成. 转录( transcription): 以一段 DNA 的遗传信息为模板,在 RNA 聚合酶作用下,合成出对应的 RNA 的过程( DNA 指导的 RNA 合成) 。 转录产物: mRNA 、rRNA、 tRNA、 小 RNA

E N D
第十四章RNA的生物合成 DNA转录 RNA的复制。 RNA DNA RNA RNA的生物合成 转录(transcription):以一段DNA的遗传信息为模板,在 RNA聚合酶作用下,合成出对应的RNA的过程(DNA 指导的RNA合成)。 转录产物:mRNA 、rRNA、 tRNA、小RNA 除某些病毒基因组RNA外,绝大多数RNA分子都来自DNA转录的产物。
第一节、DNA转录 一、转录的单位 转录通常具有一定的起点和终点,这段转录的 区域叫做转录单位。 原核生物 多顺反子 一个转录单位 真核生物 单顺反子 (一)、控制转录的起始区域—启动子 1、启动子定义:RNA聚合酶识别、结合并开始转录所必需的一段DNA序列。
2、启动子序列的测定—足迹法和DNA测序法 足迹法:即是将DNA起始转录的限制片段分离出来。加RNA聚合酶使之结合。再用DNA酶部分水解。与酶结合的部位被保护而不水解,其余部位水解成长短不同的片段,经凝胶电泳即可测出酶所结合的部位
(1)、-10序列(Pribnow框):解链区 定义:在转录起点上游大约-10处,有一个6bp的保守序列TATAAT,称Pribnow框。此段序列出现在-4到-13bp之间,解链区决定转录方向。 频度: T89 A89 T50 A65 A65 T100 作用:Pribnow框决定转录方向。酶在此部位与DNA结合形成稳定的复合物,Pribnow框中DNA序列在转录方向上解开,形成开放型起始结构,它是RNA聚合酶牢固的结合位点,是启动子的关键部位。 问题:设计一个探针能够最大数量的与E.coli启动子杂交。 (5-ATTATA-3)
-10序列对转录的效率影响 • TATAAT→AATAAT,转录效率下降,称为下降突变(down mutation)。 • TATGTT→TATATT,转录效率上升,为上升突变(up mutation)。 • 以上突变为何会影响转录效率? (前者可能由于T-A的堆集能要小于A-T的堆积能,后者可能是由于堆集能降低和氢键的减少) 据预测,Pribnow框中,一开始的TA和第6位最保守的T在结合RNA聚合酶时起十分重要的作用。
(2)、 -35序列(Sexfama box):识别区 定义:在-10序列上游一个保守序列,其中心约在-35位置,称为-35序列,此序列为RNA酶的识别区域。(只含-10序列的DNA不能转录) 碱基出现频率:T85 T83 G81 A61 C69 A52 ,其中TTG十分保守。 功能:它是原核RNA聚合酶全酶依靠σ因子的初始识别位点。-35序列的核苷酸结构,在很大程度上决定了启动子的强度,RNA聚合酶易识别强的启动子。 -35序列提供RNA聚合酶识别信号, -10序列有助于DNA局部双链解开, -35和-10序列的距离是稳定的,此与RNA pol的结构有关
(3)、转录起始位点(I) 第一个碱基在原核中常为A或G,而且位置固定
真核生物Ⅱ类基因的启动子和调控区 TATA框 核心元件 启始子(initiator,Inr):一般由PY2CAPY5构成, 位于-3~+5 ,可能提供RNA pol Ⅱ识别。 CAAT box、 Ⅱ类启动子 上游元件组成 GC box Oct .增强子(enhomcer) 远端调控区 减弱子(dehancer) 静息子(sisencer) 上游激活序列(upstream activating seguences UASs)
真核生物RNA聚合酶Ⅱ的启动子 (1)、TATA框(Hogness框):解链区 定义:中心在-25至-30,长度7bp左右。相当于原核的-10序 列。此序列功能使DNA双链解开 碱基频率:T82 A97 A85 A63 (T37 )A83 A50(T37 )(全为A- T,少数含有一个G-C对)。 功能:使DNA双链解开,并决定转录的起点位置,失去TATA框,转录将可能在许多位点上开始。 TATA框的改变或缺失,直接影响DNA与酶的结合程度,会使转录起始点偏移,因此,TATA是绝大多数真核基因正确表达所必需的。
真核生物RNA聚合酶Ⅱ的启动子 (2)、CAAT框: 聚合酶识别结合区 中心在-75处,9bp,共有序列GGT(G)CAATCT 功能:与RNA聚合酶结合。 (3)、GC框: 在CAAT框上游,序列GGGCGG,与某些转录因子结合。 CAAT和GC框均为上游序列,对转录的起始频率有较大影响。 (4)增强子 其特点是: ① 具有远距离效应。② 无方向性。③ 顺式调节。 ④ 无物种和基因的特异性。⑤ 具有组织的特异性。 ⑥ 有相位性。其作用和DNA的构象有关。 ⑦ 有的增强子可以对外部信号产生反应。
启动子比较 真核生物RNA聚合酶Ⅱ的启动子和原核生物RNA聚合酶启动子
转录因子 RNA聚合酶在进行转录时,常需要一些辅助因子(蛋白质)参与作用,此类蛋白质统称为转录因子。 人类Ⅱ型启动子的转录因子 因子 分子量 功能 RNAPolⅡ ≥10K 依赖模板合成RNA TFⅡA 12,19,35K 稳定TFⅡD和DNA的结合,激活TBP亚基 TFⅡB 33K 结合模板链(-10~+10),起始PolⅡ结合,和TFⅡE/F 相互作用 TFⅡD (TBP,30K) TBP亚基识别TATA,将聚合酶组入复合体中,TAFs识别 特殊启动子 TFⅡE 34K(β) 结合在PolⅡ的前部,使复合体的保护区延伸到下游 57K(α) TFⅡF 38, 74K 大亚基具解旋酶活性(RAP74),小亚基和PolⅡ结合, 介导其加入复合体 TFⅡH 具激酶活性,可以磷酸化PolⅡC端的CTD,使PolⅡ逸出,延伸
真核生物转录起始复合物的组装 启动子TATA盒+ TFⅡD +D-A复合物(TFⅡD+ TFⅡA) + TFⅡB +(RNA聚合酶+ TFⅡF)复合物 +TFⅡE 、TFⅡJ +TFⅡH(解旋酶、蛋白激酶) DNA解旋、RNA聚合酶C-端Ser磷酸化 从起始点向下游移动
真核生物转录起始复合物的组装 TFⅡD TFⅡB TFⅡA TFⅡF RNA聚合酶 TFⅡE 、 TFⅡJ TFⅡH
试解释在真核基因-50位点插入一个5bp的DNA序列比插入一个10bp的DNA序列对RNA聚合酶2的转录起始速率降低的程度影响更大 RNA聚合酶2的启动子除了-27位的(TATA框)外还有-50——-100之间的序列,插入一个10bp序列正 好为一个螺距,和原来的转录因子结合的位点还在 侧,插入一个5bp序列(螺旋半圈),和原来的转录 因子结合的位点在相反一侧
上游控制元件 核心元件 RNA聚合酶I类启动子 该类启动子最为简单。I类启动子控制的基因只能是rRNA基因。人类细胞中rRNA启动子由两部分组成,① 核心元件,位于转录起始部位前后-45至+20之间,负责转录的起始。② 上游控制元件(upstream control element, UCE),位于上游-180至-107之间,作用是增加核心元件转录起始的效率。这两个启动子元件的序列85%左右相同,其碱基组成的特点是富含G-C配对。
RNA聚合酶I类转录因子 • 和II类因子相比,I类因子要简单得多,只有SL1和上游结合因子(UBF)两种。 • ★ SL1 • SL1由TATA盒结合蛋白(TBP)和三种TAF组成 • (TAFI110,TAFI63和TAFI48)。 • SL1自身并不与rRNA启动子结合,但它能够扩大和增强 • RNA聚合酶I与上游控制元件的结合。 • ③ SL1有种属的特异性。 • ★ UBF:UAF能够激活完整的启动子或核心元件,并在上游控制元件的协助下介导激活。 • UBF和SL1在刺激转录的过程中有协同作用。
RNA聚合酶I I I类启动子 5sRNA tRNA SnRNA
(二)、 终止子和终止因子 1、终止子定义: 终止子:提供转录终止信号的一段DNA序列。 终止因子:协助RNA聚合酶识别终止子的蛋白质辅助因子。 2、原核生物的终止因子(terminator factors) (1).ρ蛋白:这是一种六聚体的蛋白质,亚基的分子量为50kD。该蛋白因子能识别终止信号,并能与RNA紧密结合,导致RNA的释放。 (2). nusA蛋白:为一分子量69kD的酸性蛋白,它能与RNA及RNA聚合酶相结合,在终止部位使两者被释放。
3、大肠杆菌中的两类终止子 所有原核生物的终止子在终止点之前都有一个回文结构,它转录出来的RNA可以形成一个颈环式的发荚结构。 (1)、不依赖于ρ的终止子(简单终止子) 简单终止子除具有发夹结构外,在终止点前有一寡聚U序列,回文对称区通常有一段富含GC的序列。 寡聚U序列可能提供信号使RNA聚合酶脱离模板。 (2)、依赖ρ的终止子 依赖ρ的终止子,必需在ρ因子存在时,才发生终止作用。终止点前无寡聚U序列,回文对称区不富含GC。 ρ因子是55KD的蛋白质,可水解三磷酸核苷。
(1)、不依赖于ρ的终止子(简单终止子) 不依赖于ρ因子的终止子(强终止子)的结构特点 (1) 有回文结构存在; (2) 茎的区域富内含G-C; (3)强终止子3′端上有6个U; 产物RNA形成茎-环结构是终止的关键,茎环结构使转录终止的机理: • 使RNA聚合酶变构,转录停顿; • 使转录复合物趋于解离,RNA产物释放 ☆ RNA pol变构;RNA局部双链影响DNA-RNA杂化双链 ☆ 产物出现一连串U, U :dA配对最不稳定,有利于RNA 从模板脱落。
(2)依赖ρ因子(rho factor)的终止: ρ因子是ρ基因编码的蛋白质,是一种酶,它具有ATPase的活性和解链酶的活性,在水解ATP的情况下,它沿着5′→3′方向转录物的3′端前进,直到遇到暂停在终止点位置的RNA聚合酶。随后ρ因子通过解链酶的活性解开转录泡(transcription bubble)上的RNA/DNA形成的杂交 双螺旋,使RNA转录物得到释放, 从而终止转录。 ρ因子
4、抗终止作用 通读往往发生在强启动子、弱终止子的基因上。 抗终止作用常见于某些噬菌体的时序控制。早期基因于后基因之间以终止子相隔开,通过抗终止作用可以打开后基因的表达。 λ噬菌体前早期(immediate early)基因的产物N蛋白就是一种抗终止因子。它与RNA聚合酶作用使其在左右两个终止子处发生通读,从而表达晚早期(delayed early)基因。晚早期基因的产物Q蛋白也是一种抗终止因子,它能使晚早期基因得以表达。
确定有义链 J.Marmur和Doty Spiegelnan区分有义链。采用枯草杆菌的SP8噬菌体为材料,SP8 DNA 双链有“ 轻”、“ 重”差异明显。
原核生物的聚合酶 由五个亚基组成
E.coli RNA聚合酶(原核) E.coli和其它原核细胞只有一种RNA聚合酶,合成各种RNA(mRNA、tRNA、rRNA) 1、σ亚基的功能:无σ亚基的酶叫核心酶,核心酶只能使已开始合成的RNA链延长,而不具备起始合成活性,加入σ亚基后,全酶才具有起始合成RNA的能力,因此,σ亚基称为起始因子。核心酶在DNA上滑动,σ亚基能增加酶与DNA启动子的结合常数,增加停留时间,使聚合酶迅速找到启动子并与之结合,σ亚基本身无催化活性。不同的σ因子识别不同的启动子,从而表达不同的基因。不同的原核生物,都具有相同的核心酶,但σ亚基有差别,这决定了原核基因表达的选择性。
E.coli RNA聚合酶(原核) 2、β亚基:是酶和底物结合的部位和催化位点。利福平(rifampicin)和利福霉素(rifamycin)通过结合β亚基,对全酶有强烈的抑制作用,抑制RNA合成的起始,但不抑制其延长。β亚基基因rpoB突变可使细菌对利福平有抗性。 3、β’亚基:其作用是与模板DNA相结合。β’亚基的碱性较强,适于与模板DNA相结合。肝素是一种多价阴离子,能和β‘亚基结合,从而抑制DNA与RNA聚合酶结合,进一步抑制转录作用。 4、α亚基:功能现尚未知。然而,当噬菌体T4感染E.coli时,其α亚基即在精氨酸上被ADP-核糖化。这种修饰作用使该RNA聚合酶全酶对其原先识别的启动子的亲和力减弱。所以有人认为,α亚基的功能可能参与全酶和启动子的牢固结合。
细菌RNA聚合酶特点 1. 聚合速率慢,30-85NTP/秒 2. 缺乏外切酶活性,无校对功能,错误率10-6 3. 不同的σ因子识别不同的启动子 4. 可被药物抑制(利福平) 人的RNA聚合酶对利福平不敏感。利用此特点可研制杀菌药物
纯的RNA聚合酶,在离体条件下可转录双链DNA,但在体内,DNA的两条链中只有一条可用于转录,这可能是由于RNA聚合酶在分离时丢失了σ亚基引起的或者DNA提取过程中因断裂失去了控制序列。纯的RNA聚合酶,在离体条件下可转录双链DNA,但在体内,DNA的两条链中只有一条可用于转录,这可能是由于RNA聚合酶在分离时丢失了σ亚基引起的或者DNA提取过程中因断裂失去了控制序列。 解旋和重新螺旋化也是RNA聚合酶的内在特性,在酶的前端解螺旋,在后端以相反方向重新螺旋化,活体状况中,可能还有其它酶活性来帮助调整DNA的拓扑学性质。 37℃时,RNA聚合酶的聚合速度可达50个核苷酸/秒 与蛋白质翻译速率15个氨基酸/秒相仿 但是慢于DNA复制的速率800bp /秒
原核生物RNA pol执行多功能 (1) 识别DNA双链上的启动子; (2) 使DNA变性在启动子处解旋成单链; (3) 通过阅读启动子序列,RNA pol确定它 自己的转录方向和模板链。 (4) 最后当它达到终止子时,通过识别停止转录。