Download

1 / 25
320 likes | 1.09k Views
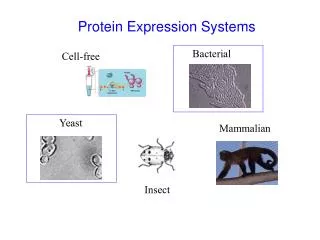
RheoSwitch ® Mammalian Inducible Expression System. (Precise Control of Gene Expression in Mammalian cells). 민지영. Vectors: pNEBR-R1 Regulator Plasmid (500 μg/ml) 20 μg

E N D
RheoSwitch® Mammalian Inducible Expression System (Precise Control of Gene Expression in Mammalian cells) 민지영
Vectors: • pNEBR-R1 Regulator Plasmid (500 μg/ml) 20 μg • Expresses an engineered nuclear receptor heterodimer, consisting of two proteins, RheoReceptor-1 and RheoActivator. The two proteins constitute the holoreceptor and regulate transcription of genes cloned into the expression vector, pNEBR-X1.The two proteins are expressed from the constitutive ubiquitin UbC and UbB promoters, respectively. The plasmid also contains the neomycin resistance gene under control of the SV40 early promoter for the generation of stable cell lines. • 2. pNEBR-X1Hygro Vector (500 μg/ml) 20 μg • Used to clone the gene of interest. The plasmid, has five copies of the GAL4 response element (5XRE) upstream of a TATA box and a short leader sequence followed by multiple cloning sites and an SV40 polyA signal. The plasmid also contains the hygromycin resistance gene under control of the thymidine kinase promoter for generation of stable cell lines. • 3. pNEBR-X1GLuc Control Plasmid (500 μg/ml) 20 μg • Used as a positive control for expression analysis. This plasmid expresses secreted Gaussia luciferase.
1. pNEBR-R1 plasmid map. Fig1. Expresses an engineered nuclear receptor heterodimer- RheoReceptor-1, RheoActivator. The two proteins constitute the holoreceptor and regulate transcription of genes cloned into the expression vector, pNEBR-X1. The two proteins are expressed from the constitutive UbC and UbB promoters, respectively. The plasmid also contains the neomycin resistance gene under control of the SV40 early promoter for the generation of stable cell lines.
2. pNEBR-X1Hygro plasmid map. Fig2. Used to clone the gene of interest, it has five copies of the GAL4 response element (5XRE) upstream of a TATA box and a short leader sequence followed by multiple cloning sites and an SV40 polyA signal. Only unique restriction enzyme sites are shown.
2.1 pNEBR-X1Hygro multiple cloning sites (MCS). Fig 2-1. To minimize uninduced expression, a short sequence containing the TATA box separates the end of the 5XRE from the predicted transcription start. Only unique restriction enzyme sites are shown.
Inducer: RheoSwitch® Ligand RSL1 (5 mM in DMSO) 50 μl Sequencing Primer: RheoSwitch R-X1 Sequencing Primer 5´ (GGGTATATAATGGGGGC) 3´ 200 pmol This primer can be used to confirm correct insertion of gene to be expressed in pNEBR-X1.Supplied as a lyophilized triethylammonium salt. Luciferase Assay Reagents: GLuc Substrate (100X) 50 μl GLuc Assay Buffer 5 ml
Fig3. • RheoReceptor : LBD of an insect EcR nuclear recepter • to fused to the yeast GAL4 (DBD) • RheoActivator : Insect/mammalian RXR hybrid LBD • fused to the viral activation domain VP16
< Example protocol > Day1 : NIH 3T3 cells in a 24 well plate 2.5*104 cells (50-70%) per well in 0.5ml DMEM
Gene of interest pNEBR-R1 pNEBR-X1 Hygro RSL1 (in DMSO) DMSO ( as a control) Day2 : 600ng pNEBR-R1 (100ng/ml) & 2.4ug pNEBR-X1 Gluc (400ng/ml) transfection Add RSL1 ligand to a final concentration of 500nM Mock : DMSO (total volume의 0.1% (v/v)가 넘지 않도록 한다.
Gene of interest pNEBR-X1 Hygro pNEBR-R1 RSL1 ligand 2. 만들어진 cell line에 pNEBR-X1 (gene)-Hygro transfection (Hygromycin으로 selection) 3. RSL1 ligand treatment Inducible Expression 1. Cell line 만들기 (RheoSwitch receptor Stable cell line)
Fig4. Luciferase expression in response to increasing RSL1 concentrations. NIH 3T3 cells co-transfected with pNEBR-R1 and pNEBR-X1 expressing firefly luciferase were induced with the indicated RSL1 concentrations. Expression was measured 48 hours post-induction.
Fig5. NIH 3T3 (A) and RheoSwitch HEK293-A7 (B) (NEB #C2003) cells were transfected with pNEBR-X1Hygro encoding a HA-tagged protein using TransPass D2 (NEB #M2554). NIH 3T3 cells were also co-transfected with pNEBR-R1. RSL1 was added to transfected cells 16 hours later, and cells were harvested 24 hours after induction (Note: DMSO is added as a control for 0 nM RSL1). Harvested cells were lysed, and equivalent amounts of protein (as determined by Coomassie Stain) were electrophoresed and transferred to nitrocellulose. An immunoblot assay was performed on the filter using anti-HA as primary antibody and detected using anti-mouse secondary antibody and reagents. A longer exposure of the HEK293-A7 immunoblot is shown (C) to illustrate that no protein is detected in the absence of RSL1.
Fig 6. Induction time course. NIH 3T3 cells co-transfected with pNEBR-R1 and pNEBR-X1GLuc control plasmid were induced with RSL1. Gaussia luciferase activity was assayed between 1 and 24 hours post-induction.
Fig 7. Rapid ON:OFF switch response to two cycles of addition and removal of RSL1. NIH 3T3 cells were co-transfected with pNEBR-R1 and pNEBR-X1 expressing a destabilized firefly luciferase and induced with RSL1 followed by removal of RSL1 at 24 hours. A second cycle of addition/removal of RSL1 results in a new ON:OFF switch response. To remove RSL1 and turn off expression, cells were washed and RSL1-free media was added at the times indicated. The blue line represents induced expression. The red line represents uninduced expression.
Several regulatory sequences may be found in the 5' UTR: • Binding sites for proteins, that may effect the mRNA's stability or translation, for example iron responsive elements, which occur in the 5' UTRs (and 3' UTRs) of a small number of eukaryotic mRNAs that regulate gene expression in response to iron. • Regulatory elements that do not depend on proteins, such as riboswitches. • Sequences that promote the initiation of translation.
The five prime untranslated region (5' UTR), also known as the leader sequence, is a particular section of messenger RNA (mRNA) and the DNA that codes for it. It starts at the +1 position (where transcription begins) and ends just before the start codon (usually AUG) of the coding region. It usually contains a ribosome binding site (RBS), in bacteria also known as the Shine Dalgarno sequence (AGGAGGU). The 5' UTR may be a hundred or more nucleotides long, and the 3' UTR may be even longer (up to several kilobases in length) An mRNA molecule codes for a protein through translation. The mRNA also contains regions that are not translated: in eukaryotes this includes the 5' untranslated region, 3' untranslated region, 5' cap and poly-A tail. In prokaryotic mRNA the 5' UTR is normally short. Some viruses and cellular genes have unusual long structured 5' UTRs which may have roles in gene expression.
LBD of an insect EcR nuclear receptor Pulses of 20-hydroxyecdysone occur during insect development, whereupon this hormone binds to the ecdysone receptor, a ligand-activated transcription factor found in the nuclei of insect cells. This in turn leads to the activation of many other genes, as evidenced by chromosomal puffing at over a hundred sites. Ultimately the activation cascade causes physiological changes that result in ecdysis (moulting). Namely an N-terminal transcriptional activation domain (A/B domain), a DNA-binding domain (C domain, highly conserved between receptors), a linker region (D region), a ligand-binding domain (E domain, moderately conserved), and in some cases a distinct C-terminal extension (F-domain). The DNA-binding domains of EcR and USP recognize specific short sequences in DNA, and mediate the binding of the heterodimer to these ecdysone response elements (ECREs) in the promoters of ecdysone-responsive genes.
GAL4, DNA-binding domain The nuclear protein GAL4 is a positive regulator of gene expression for the galactose-induced genes such as GAL1, GAL2, GAL7, GAL10, and MEL1. These genes encode enzymes which convert galactose to glucose. GAL4 recognizes a 17 base-pair long sequence in the upstream activating sequence (uas-g) of these genes, (5'-cggrnnrcynyncnccg-3'). GAL4 binds to the DNA as a homodimer, most likely forming a complex with GAL11.This protein contains a fungal Zn(2)-Cys(6) binuclear cluster domain. Many transcriptional activator proteins possess such a domain in which six conserved cysteine residues bind to two zinc ions known as a binuclear zinc cluster. This cysteine-rich region binds to the DNA in a zinc-dependent fashion. While GAL4 from the organism Saccharomyces cerevisiae contains such a domain, it binds two Cadmium (Cd) ions rather than Zinc ions. The metal-dependent DNA-binding domain of GAL4 is shown above. It is 43 amino acid residues long. It contains two right-handed alpha-helices shown here in red. The six conserved cysteine residues are shown in blue, and the two cadmium ions are shown in silver.
Five HSV genes: a4--ICP4, a0--ICP0, a27--ICP27/UL 54, a22--ICP22/US1, and a47--ICP47/US12 are expressed and function the earliest stages of the productive infection cycle. This stage of infection is termed the "immediate-early" or "a" phase of gene expression, and is mediated by the action of a-TIF through its interaction with cellular transcription factors at specific enhancer elements associated with the individual a-transcript promoters. Proteins encoded by the a4, a0, and a27 transcripts act to activate viral gene expression at the level of transcription, or at least, mRNA expression. They functionally interact to form nuclear complexes with viral genomes. Surprisingly, only two (a4 and a27) have extensive areas of sequence similarity among a large number of alpha-herpesviruses, and only amino acid sequences in a27 appear to be extensively conserved among the more distantly related beta- and gamma-herpesviruses. Both of the two other a proteins, a22 and a47 are dispensable for virus replication in many types of cultured cells, but a22 is required for HSV replication in others and may have a role in maintaining the virus's ability to replicate in a broad range of cells in the host -- perhaps by providing some types of cells with the capacity to express of a group of late transcripts. The a47 protein appears to have a role in modulating host response to infection by specifically interfering with the presentation of viral antigens on the surface of infected cells.
Early gene expression Activation of the host cell transcriptional machinery by the action of a gene products, results in the expression of the early or b genes. The promoters for such genes (exemplified by the thymidine kinase transcript promoter) have served as models for "typical" eukaryotic promoters. Seven of these are necessary and sufficient for viral DNA replication under all conditions: DNA polymerase (UL30), DNA binding proteins (UL42 and UL29 or ICP8), ORI binding protein (UL9), and the helicase/primase complex (UL5, 8, and 52). When sufficient levels of these proteins have accumulated within the infected cell, viral DNA replication ensues. Other early proteins are involved in increasing the deoxyribonucleotide pools of the infected cells, while still others appear to function as repair enzymes for the newly synthesized viral genomes. These accessory proteins are "non-essential" for virus replication in that cellular products can substitute for their function in one or another cell type or upon replication of previously quiescent cells; however, disruptions of such genes often have a profound effect upon viral pathogenesis, and/or ability to replicate in specific cells.
Genome replication and late gene expression The vegetative replication of viral DNA represents a critical and central event in the viral replication cycle. High levels of DNA replication irreversibly commit a cell to producing virus, which eventually results in cell destruction. DNA replication also has a significant influence on viral gene expression. Early expression is significantly reduced or shut off following the start of DNA replication, while late genes begin to be expressed at high levels. Transcripts expressing late genes can be divided into two subclasses: "leaky-late" (bg) and "strict" late (g). Promoters controlling expression of both classes are similar in that both have elements near the transcription start site (cap site) which are required for promoter activity, but the location of other elements can differ. The bg transcripts are expressed at low levels prior to DNA replication, but reach maximum expression after viral DNA replication has been initiated. In contrast, g transcripts are difficult to detect at all until the onset of viral DNA replication.