Download
1 / 1
10 likes | 142 Views
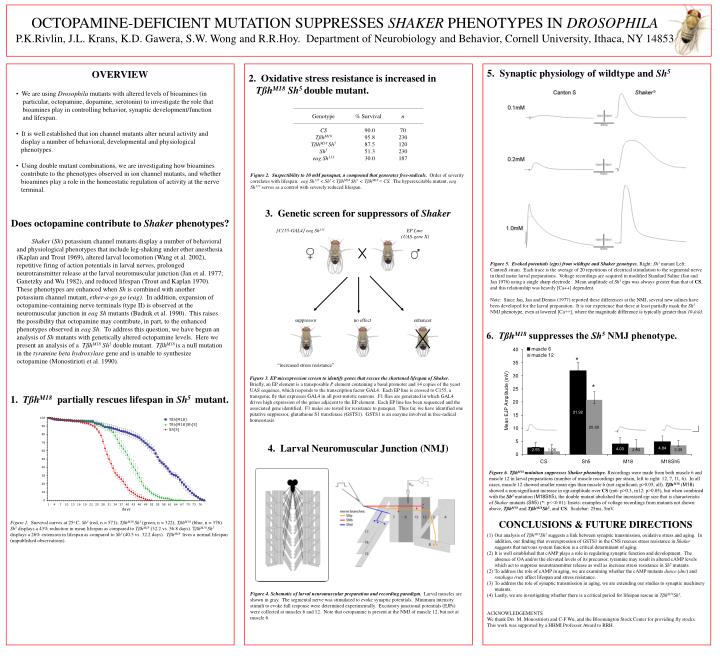
[C155-GAL4] eag Sh 133. EP Line (UAS-gene X). Genotype CS Tßh M18 Tßh M18 Sh 5 Sh 5 eag Sh 133. % Survival 90.0 95.8 87.5 51.3 30.0. n 70 230 120 230 187. suppressor. no effect. enhancer. “increased stress resistance”.

E N D
[C155-GAL4] eag Sh133 EP Line (UAS-gene X) Genotype CS TßhM18 TßhM18 Sh5 Sh5 eag Sh133 % Survival 90.0 95.8 87.5 51.3 30.0 n 70 230 120 230 187 suppressor no effect enhancer “increased stress resistance” OCTOPAMINE-DEFICIENT MUTATION SUPPRESSES SHAKER PHENOTYPES IN DROSOPHILA P.K.Rivlin, J.L. Krans, K.D. Gawera, S.W. Wong and R.R.Hoy. Department of Neurobiology and Behavior, Cornell University, Ithaca, NY 14853 5. Synaptic physiology of wildtype and Sh5 OVERVIEW Oxidative stress resistance is increased in TßhM18 Sh5 double mutant. • We are using Drosophila mutants with altered levels of bioamines (in • particular, octopamine, dopamine, serotonin) to investigate the role that • bioamines play in controlling behavior, synaptic development/function • and lifespan. • It is well established that ion channel mutants alter neural activity and display a number of behavioral, developmental and physiological phenotypes. • Using double mutant combinations, we are investigating how bioamines contribute to the phenotypes observed in ion channel mutants, and whether bioamines play a role in the homeostatic regulation of activity at the nerve terminal. Figure 2. Suspectibility to 10 mM paraquat, a compound that generates free-radicals. Order of severity correlates with lifespan: eag Sh133< Sh5 < TßhM18 Sh5 < TßhM18 ≈ CS. The hyperexcitable mutant, eag Sh133serves as a control with severely reduced lifespan. 3. Genetic screen for suppressors of Shaker Does octopamine contribute to Shaker phenotypes? Shaker (Sh) potassium channel mutants display a number of behavioral and physiological phenotypes that include leg-shaking under ether anesthesia (Kaplan and Trout 1969), altered larval locomotion (Wang et al. 2002), repetitive firing of action potentials in larval nerves, prolonged neurotransmitter release at the larval neuromuscular junction (Jan et al. 1977; Ganetzky and Wu 1982), and reduced lifespan (Trout and Kaplan 1970). These phenotypes are enhanced when Sh is combined with another potassium channel mutant, ether-a-go go (eag). In addition, expansion of octopamine-containing nerve terminals (type II) is observed at the neuromuscular junction in eag Sh mutants (Budnik et al. 1990). This raises the possibility that octopamine may contribute, in part, to the enhanced phenotypes observed in eag Sh. To address this question, we have begun an analysis of Sh mutants with genetically altered octopamine levels. Here we present an analysis of a TßhM18 Sh5 double mutant. TßhM18 is a null mutation in the tyramine beta hydroxylase gene and is unable to synthesize octopamine (Monostirioti et al. 1990). Figure 5. Evoked potentials (ejps) from wildtype and Shaker genotypes.Right: Sh5 mutant Left: CantonS strain. Each trace is the average of 20 repetitions of electrical stimulation to the segmental nerve in third instar larval preparations. Voltage recordings are acquired in modified Standard Saline (Jan and Jan 1976) using a single sharp electrode . Mean amplitude of Sh5ejps was always greater than that of CS, and this relationship was heavily [Ca++] dependent. Note: Since Jan, Jan and Dennis (1977) reported these differences at the NMJ, several new salines have been developed for the larval preparation. It is our experience that these at least partially mask the Sh5NMJphenotype, even at lowered [Ca++], where the magnitude difference is typically greater than 10-fold. 6. TßhM18suppresses the Sh5NMJ phenotype. Figure 3. EP misexpression screen to identify genes that rescue the shortened lifespan of Shaker. Briefly, an EP element is a transposable P element containing a basal promoter and 14 copies of the yeast UAS sequence, which responds to the transcription factor GAL4. Each EP line is crossed to C155, a transgenic fly that expresses GAL4 in all post-mitotic neurons. F1 flies are generated in which GAL4 drives high expression of the genes adjacent to the EP element. Each EP line has been sequenced and the associated gene identified. F1 males are tested for resistance to paraquat. Thus far, we have identified one putative suppressor, glutathione S1 transferase (GSTS1). GSTS1 is an enzyme involved in free-radical homeostasis. 1. TßhM18partially rescues lifespan in Sh5 mutant. 4. Larval Neuromuscular Junction (NMJ) Figure 6. TßhM18 mutation suppresses Shaker phenotype.Recordings were made from both muscle 6 and muscle 12 in larval preparations (number of muscle recordings per strain, left to right: 12, 7, 11, 6). In all cases, muscle 12 showed smaller mean ejps than muscle 6 (not significant; p>0.05, all). TßhM18 (M18) showed a non-significant increase in ejp amplitude over CS (m6: p>0.3, m12: p>0.05), but when combined with the Sh5 mutation (M18Sh5), the double mutant abolished the increased ejp size that is characteristic of Shaker mutants (Sh5) (*: p<<0.01). Insets: examples of voltage recordings from mutants not shown above, TßhM18 and TßhM18Sh5, and CS. Scalebar: 25ms, 5mV. CONCLUSIONS & FUTURE DIRECTIONS Figure 1. Survival curves at 25o C. Sh5 (red, n = 571); TßhM18 Sh5(green, n = 322); TßhM18 (blue, n = 376). Sh5 displays a 43% reduction in mean lifespan as compared to TßhM18 (32.2 vs. 56.8 days). TßhM18 Sh5 displays a 26% extension in lifespan as compared to Sh5 (40.5 vs. 32.2 days). TßhM18 lives a normal lifespan (unpublished observations). Our analysis of TßhM18Sh5 suggests a link between synaptic transmission, oxidative stress and aging. In addition, our finding that overexpression of GSTS1 in the CNS rescues stress resistance in Shaker suggests that nervous system function is a critical determinant of aging. It is well established that cAMP plays a role in regulating synaptic function and development. The absence of OA and/or the elevated levels of its precursor, tyramine may result in altered cAMP levels which act to suppress neurotransmitter release as well as increase stress resistance in Sh5 mutants. To address the role of cAMP in aging, we are examining whether the cAMP mutants dunce (dnc) and rutabaga (rut) affect lifespan and stress resistance. To address the role of synaptic transmission in aging, we are extending our studies to synaptic machinery mutants. Lastly, we are investigating whether there is a critical period for lifespan rescue in TßhM18Sh5. ACKNOWLEDGEMENTS We thank Drs. M. Monostirioti and C-F Wu, and the Bloomington Stock Center for providing fly stocks. This work was supported by a HHMI Professor Award to RRH. Figure 4. Schematic of larval neuromuscular preparation and recording paradigm. Larval muscles are shown in gray. The segmental nerve was stimulated to evoke synaptic potentials. Minimum intensity stimuli to evoke full response were determined experimentally. Excitatory junctional potentials (EJPs) were collected at muscles 6 and 12. Note that octopamine is present at the NMJ of muscle 12, but not at muscle 6.