Download

1 / 27
310 likes | 536 Views
Biological Bases of Behaviour. Lecture 4: Synaptic Transmission. Learning Outcomes. At the end of this lecture you should be able to: 1 . Explain what is meant by a 'reflex arc'. 2 . Describe how research on reflexes has contributed to our understanding of synaptic events.

E N D
Biological Bases of Behaviour. Lecture 4: Synaptic Transmission.
Learning Outcomes. • At the end of this lecture you should be able to: • 1. Explain what is meant by a 'reflex arc'. • 2. Describe how research on reflexes has contributed to our understanding of synaptic events. • 3. Describe how behaviour can be related to synaptic integration. • 4. Explain the major chemical events at the synapse.
Reflexes. • In the late 1800's Cajal demonstrated that neurons do not physically touch one another - they are separated by a tiny gap called a synapse. • From experiments assessing reflexes (automatic responses to stimuli) Sherrington (1906) proposed that neurons communicated with one another via the synapse. • In a typical leg-flexing reflex a sensory neuron excites a second neuron (interneuron) which in turn excites a motor neuron which triggers a muscle to contract. • This entire process is called a reflex arc.
Reflex Arc for Leg Flexion. Brain neuron Interneuron skin Sensory neuron Spinal column Motor neuron muscle Kalat (2001), p 53
Characteristics of the Reflex. • Sherrington observed certain properties of the reflex which convinced him that a specific process was happening at the junctions between neurons: • 1. Speed: He measured the total distance that an impulse travels from sensory receptor to spinal cord to muscle, and calculated the speed of the impulse. • He found that the speed of conduction through a reflex arc was significantly slower than that along a single axon, therefore there must be some delay at the synapses. • 2. Summation: When a weak stimulus is applied (a pinch) a reflex may not be produced, however if several small pinches are rapidly applied they trigger a reflex. • This is called temporal summation.
Summation. • Sherrington’s ideas were confirmed by Eccles (1964). • He attached stimulating electrodes to 2 axons that formed a synapse and recorded from one neuron whilst repeatedly stimulating the other. • During stimulation, brief depolarizations of the membrane potential were seen (it became more positive) such depolarizations are called excitatory postsynaptic potentials (EPSP's). • If several EPSP’s occur within a specific time then an action potential will be triggered in the postsynaptic cell (temporal summation). • Alternatively, single EPSP’s from different axons can combine to exceed the threshold of excitation of the postsynaptic cell - this is called spatial summation.
Temporal and Spatial Summation. Temporal summation (several impulses from one neuron over time) Action potential Spatial summation (impulses from several neurons at the same time) Kalat (2001), p 54
Synaptic Activation. Simultaneous EPSP’s combine spatially to exceed the threshold EPSP Temporal summation of 2 EPSP’s Resting potential 3 EPSP’s combine to exceed the threshold IPSP Kalat (2001), p 54
Characteristics of the Reflex continued. • 3. Inhibition: When stimulation is applied a flexor muscle will contract, at the same time an extensor muscle will relax, so while one muscle is being stimulated the other is being inhibited. • Sherrington realised that the second neuron (interneuron) in the reflex arc must also connect to an inhibitory motor neuron. • Eccles (1964) showed that the input from an axon not only depolarizes the postsynaptic neuron but can also hyperpolarize it (making it more negative), and thus less likely to fire. • This temporary hyperpolarization is called an inhibitory postsynaptic potential (IPSP).
Inhibitory Synapses. Brain neuron Excitatory synapse skin Interneuron Inhibitory synapse Sensory neuron Motor neuron to extensor muscle Motor neuron to flexor muscle Kalat (2001), p 55
Perceptual Experience. • The all or none law states that an action potential is either triggered or not, this gives the impression that our sensory experiences are either experienced or not. • Clearly this does not happen, e.g. muscle contractions can be weak or intensive, a spot of light can be perceived as being faint or very bright. • If the action potential is pulse-like how can it represent the intensity of a stimulus? • The answer is that a single action potential does not act as a specific unit of information, instead variable information is provided by the axon's rate of firing. • E.g a high rate of firing may single a very bright light in the retinal receptors but a low rate of firing may signal a very dim light.
Sensation and Neural Integration. • The all or none law is supplemented by the Rate law which states that the strength of a stimulus is represented by the firing rate of an axon, despite the fact that the action potential remains constant. • The rate of firing of an axon is determined by whether the sum of its connections are excitatory or inhibitory - this is referred to as neural integration. • The greater the number of EPSP's the greater the probability of an action potential, the greater the number of IPSP's the lower the likelihood of an action potential. • If a neuron is excited or inhibited, this does not necessarily mean that behaviour is either excited or inhibited. E.g if inhibitory neurons are inhibited then the result will be excitation - e.g. alcohol and prefrontal cortex.
Simple Neural Integration. Axon A stimulated = depolarization Stimulate axon A Axon B stimulated = hyperpolarization A C Both axons stimulated Excitatory synapse Inhibitory synapse B Stimulate axon B Carlson (1994) p41
The Nature of Synaptic Transmission. • Loewi(1920’s) stimulated the vagus nerve in a frogs heart causing heart rate to decrease. Fluid from this heart injected into a second, caused it to also slow down. • Stimulating the accelerator nerve (speeds up heartbeat) and injecting fluid from this heart into a second caused it to be also stimulated. • Synaptic transmission was thus shown to be chemical in nature.
Events at the Synapse. • The major events at the synapse are as follows: • 1. Synthesis: Neurotransmitters are synthesised within the terminal button by the Golgi apparatus ensuring that neurotransmitter turnover is rapid. • Peptides are synthesised within the soma, which lies far from the releasing point; so peptide turnover is much slower. • Neurotransmitters are synthesised from precursor molecules derived form the diet, e.g acetylcholine is synthesised from choline found in cauliflower and milk. • Both neurotransmitters and peptides are stored in spherical packets called synaptic vesicles.
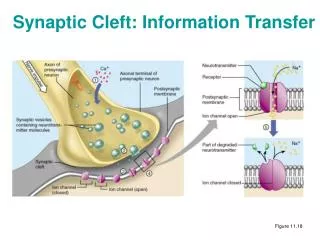
Events at the Synapse (continued). • 2. Transport: Synaptic vesicles are transported down the axon to the terminal buttons. They collect in the release zone and 'dock' against the presynaptic membrane facing the synaptic cleft. • 3. Release: The release zone contains voltage-dependent calcium channels and so when an action potential reaches the terminal button, it depolarizes the presynaptic membrane, opening the calcium channels. • The influx of calcium opens specialised channels in the membrane called fusion pores which allow the neurotransmitter stored in the vesicles to be released into the synaptic cleft. • This whole process is called exocytosis and lasts around 1-2 milliseconds.
Release of Neurotransmitters. Carlson (1994) p51 Undocked vesicle Protein molecules Docked vesicle Entry of calcium opens fusion pore Transmitter molecules leave button Fusion pore widens
Synaptic Cross-Section. vesicles Presynaptic membrane Vesicles releasing neurotransmitter Postsynaptic membrane Carlson (1994) p50
Events at the Synapse (continued). • 4. Attachment: Molecules of neurotransmitter diffuse across the synaptic cleft to the postsynaptic membrane where they attach to the binding sites of specialised protein receptors. • A neurotransmitter molecule fits into a binding site like a key in a lock so receptors only work with a specific neurotransmitter. • To complicate matters each neurotransmitter can have several different types of receptor each with different characteristics. • As neurotransmitters are common chemicals there are many natural and artificial chemicals (drugs, poisons) that mimic their effects as they also fit the same binding sites. Any chemical that attaches to a binding site is called a ligand.
Events at the Synapse (continued). • 5. Activation: When a neurotransmitter has attached to a binding site it can have several effects depending upon the nature of the receptors it has attached to. • a) Ionotropic effects: These are very rapid but short-lived. • According to North (1989) a neurotransmitter having this effect opens the ion channels within 10ms after its release and keeps them open for less than 20ms. • Such events are useful for conveying rapid information about sensory and muscular stimulation. • Ionotropic effects can be excitatory or inhibitory, the most common excitatory ionotropic transmitter is glutamate, while the most common inhibitory one is GABA.
Ionotropic Receptors. Neurotransmitter attached to binding site Ions Open ion channel Closed ion channel Carlson (1994) p54
b) Metabotropic Effects. • These effects are slower and longer-lasting as they involve a sequence of metabolic reactions relying upon a secondary messenger. • According to North (1989) they take place 30ms or more after the release of the neurotransmitter and may last minutes or hours. • In such receptors the arrival of the neurotransmitter activates G-protein and then one of two things happen:
Metabotropic Receptors. • 1. An subunit breaks away from the G-protein, binds directly with the ion channel and opens it. Carlson (1994) p55
Metabotropic Receptors (continued). • 2. An subunit breaks away from the G-protein,activates another enzyme which then produces a second messenger (e.g cyclic AMP) which in turn influences the ion channel. Carlson (1994) p55
Events at the Synapse (continued). • 6. Inactivation: Neurotransmitters are removed from the synapse in several ways: • Deactivation: An enzyme destroys the transmitter molecule, e.g acetylcholine is broken down into choline and acetate by acetylcholinesterase (AChE), the choline then returns to the presynaptic neuron where it is recombined with acetate to form more neurotransmitter. • Reuptake: Other transmitter substances detach from their receptors and are taken back to the presynaptic membrane by carrier molecules. They enter the membrane via special membrane proteins called transporters, where they are reused.
Reuptake. Transmitter molecules returned to button transporter Carlson (1994) p57
References and Bibliography • Carlson, N.R. (1994). Physiology of Behaviour. • Eccles, J.C. (1964). The Physiology of Synapses. • Kalat, J.W. (2001). Biological Psychology. • North, R.A. (1989). Neurotransmitters and their receptors: from the clone to the clinic. Seminars in the Neurosciences, 1: 81-90. • Soltesz, I., Smetters, D.K., & Mody, I. (1995). Tonic inhibition originates from synapses close to the soma. Nature, 14: 1273-1283.